֬�����л(37��ҽѧ����
2012-07-22 20:37:14 ��Դ��37��ҽѧ�� ���ߣ�37��ҽѧ�� ���ۣ�0 �����
һ��֬����������ֽ�
����֬�������г���������������£��������ֽ�ΪCO2��H2O���ͷŴ������������֬�����ǻ�����Ҫ������Դ֮һ���κͼ����ǽ���֬�����������Ծ����֯��������Ҫ��������ʽ��β-������
����(һ)֬�����β-��������
�����˹��̿ɷ�Ϊ���ת�ƣ�β-�����������Ρ�
����1.֬����Ļ
������������һ����֬����μӴ�лǰҲ��Ҫ�������ʽ��������֬����CoA����֬������ø��֬��CoA�ϳ�ø(acyl CoA synthetase)��
����
����������ɵ�֬��CoA������ǿ��������ˮ���������и��ܼ������ʻ��ã���ø����������ø����������˸����ײμӷ�Ӧ��
����֬��CoA�ϳ�ø�ֳ���ø���ֲ��ڰ����С�������Ĥ��������Ĥ�ϡ������е���ø��
�����ж���֬������������Ĥ�ϵ�ø�����֬���ᣬ����֬��CoA��Ȼ��������������ڸ��������ϳɣ���������Ĥ�ϵ�ø��ij���֬��CoA���������������β-������
����2.֬��CoA���������壺��֬����β-������øϵ������������У�������֬��CoA��������ͨ����������Ĥ��Ҫ������������ʾ���Ҫ����ת�ˣ���һ���������ⶾ��(carnitine)����3���ǣ�4�����װ������ᡣ

��������֬����CoA���ⶾ�Ӧ�����ɸ�øA��֬���ⶾ�֬���������ⶾ���3���ǻ�ͨ�����������ӡ�
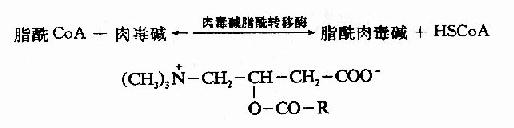
�������˷�Ӧ��øΪ�ⶾ��֬��ת��ø(carnitine acyl transferase)����������Ĥ������������д�ø��ϵͬ��ø���ֱ��Ϊ�ⶾ��֬��ת��øI���ⶾ��֬��ת��ø��ø��ʹ������֬��CoAת��Ϊ�� øA��֬�����ⶾ����߽�����������Ĥ��λ����������Ĥ�ڲ��ø����ʹ֬�����ⶾ��ת�����ⶾ���֬��CoA���ⶾ�����·������������ܣ�֬��CoA�������������ʣ���Ϊ֬����β-����øϵ�ĵ���(ͼ5��10)��
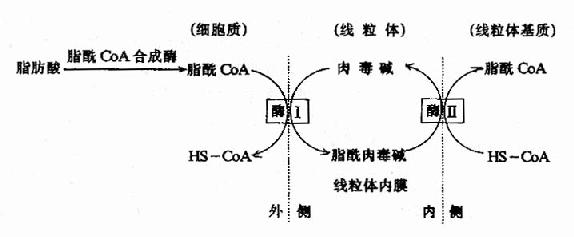
ͼ5-10���ⶾ�����֬����øAת��������ʾ��ͼ
ø��λ����������Ĥ�����ⶾ��֬��ת��ø��
ø��λ����������Ĥ�ڲ���ⶾ��֬��ת��ø
��������֬��CoA������������ٶ��ܵ��ⶾ��֬��ת��ø���ø��ĵ��ڣ�ø���ܱ�����CoA���ƣ�ø�����ȵ��� ���ơ�������CoA�Ǻϳ�֬�����ԭ�ϣ��ȵ���ͨ���յ�����CoA�Ȼ�ø�ĺϳ�ʹ������CoAŨ�����ӣ���������ø���Կ����ȵ��ض��ⶾ��֬��ת��ø ���ø���м�ӻ�ֱ���������á��������ʳʱ�ȵ��ط��ڼ��٣��ⶾ��֬��ת��ø���ø��������ߣ�ת�Ƶij���֬��������������������ܡ�
����3.β-�����ķ�Ӧ���̣�֬��CoA������������н���β����Ҫ�����IJ���Ӧ�������⡢��ˮ�����������⣬����һ��������CoA��һ��������̼���µ�֬��CoA��
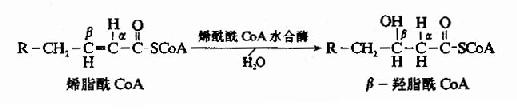
������һ������(dehydrogenation)��Ӧ��֬��CoA����ø�������ΪFAD��֬��CoA��α��β̼ԭ���ϸ���ȥһ����ԭ�����ɾ��з�ʽ˫����α��β-ϩ֬������øA��

�����ڶ�����ˮ��hydration)��Ӧ��ϩ��CoAˮ��ø�������ɾ���L-���͵�β-��֬��CoA��
�������������ⷴӦ����β����֬����CoA����ø����øΪNAD+�����£�β-��֬����CoA��������βͪ֬��CoA��
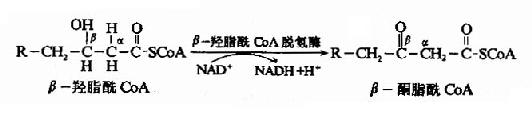
�������IJ���⣨thiolysis)��Ӧ��β��ͪ���ø����β��ͪ����CoA��α��β̼ԭ��֮�����������һ���Ӹ�øA��������CoA��һ��������̼ԭ�ӵ�֬��CoA��
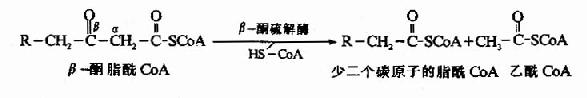
���������IJ���Ӧ��TCAѭ�����������ᾭ�Ӻ����ᡢƻ�������ɲ�������Ĺ������ƣ�ֻ��β-�����ĵ��IJ���Ӧ����⣬�������������һ����Ӧ��������CoA�������������ᡣ
��������֬��CoA������һ��ѭ����̼����������̼ԭ�ӣ�����һ��������CoA������ظ������ѭ�����ͻ�����������CoA��
�������������Կ���֬�����β-�������̾��������ص㡣����Ҫ��֬��������֬��CoA������һ�����ܹ��̡� �С�����֬����������ֱ�����������壬������֬��CoA��Ҫ�ⶾ��ת�ˡ�β-������Ӧ���������ڽ��У����û��������ĺ�ϸ����������֬���ṩ�ܡ� β-������������FADH2��NADH+H+���ɣ���Щ��Ҫ�����������ݸ�������ˮ����Ҫ���μӣ�����CoA������Ҳ��Ҫ������ˣ�β-�����Ǿ��������� ���̡�
����֬����β-�������������̿�����ͼ(ͼ5��11)��ʾ��

ͼ5-11��֬����β������Ӧ����
����(��)֬����β-��������������
����֬����β-����������֬����ֽ����Ҫ;����֬�����������Թ�Ӧ��������Ҫ�Ĵ�����������ʮ�˸�̼ԭ�ӵı���֬����Ӳ֬��Ϊ������β-�������ܷ�ӦΪ��
����CH3(CH2)15COSCoA+8NAD++*CoASH+8H2O——→9CH3COSCoA+8FADH2+8NADH+8H+����
����8����FADH2�ṩ8×2=16����ATP��8����NADH+H+�ṩ8×3=24����ATP��9�������� CoA��ȫ�����ṩ9×12=108������ATP�����һ�˷���Ӳ֬����ȫ��������CO2��H2O�����ṩ148�˷���ATP��Ӳ֬��Ļ��������2�� ����ATP������һ�˷���Ӳ֬����ȫ�����ɾ�����146�˷���ATP��һ�˷�����������ȫ����������38����ATP�����˷�������������̼ԭ������һ�˷� ��Ӳ֬����ͬ��ǰ�߿��ṩ114�˷���ATP�����߿��ṩ146�˷���ATP���ɼ���̼ԭ������ͬ�������֬�������ṩ�����������֬��������ʱ�ͷų��� ������Լ��40%Ϊ�������úϳɸ��ܻ��������60%���ȵ���ʽ�ͳ�����Ч��Ϊ40%��˵�������ܺ���Ч������֬�����������ṩ��������
����֬����β-����Ҳ��֬����ĸ�����̣���������Ҫ��֬�������ij��̲�ͬ��ͨ��β-�����ɽ�����֬�������ɳ������˵�֬���ᣬ�������л���衣
����֬����β-�������������ɵ�����CoA��һ��ʮ����Ҫ���м仯�������CoA���ܽ���������ѭ�����������⣬����������Ҫ������ϳɵ�ԭ�ϣ���ͪ�塢���̴�����̴������
����(��)֬���������������ʽ
����1.����������������ں���ʳ�к�������������̼ԭ��֬���ᣬ����β-��������������CoA�����һ���ӱ� ��CoA��ijЩ���������������ᡢ��������հ���ķֽ��л�������б���CoA���ɣ���֭�����ɹ��������������CoA������CoA�����Ȼ���Ӧ�ͷ����� ���ţ���ת������������CoA���ɽ�һ�������ֽ⣬Ҳ�ɾ����������������ǣ���Ӧ���̼���ͼ��
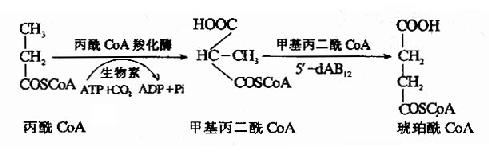
������������CoA��λø�ĸ�ø��5′����������B12(5′dAB12)��ά����B12ȱ����5′ ��dAB12�����ϰ���Ӱ���λø���ԣ�ʹ��������CoA�ѻ��������һ�����������CoA��ȥ��øA�����ɼ�����������Ѫ�м������Ậ������ (��������Ѫ֢)�����������ų�����(24Сʱ�ų�������4mgʱ��Ϊ����������֢)����һ�������������CoAŨ�����ߣ��ɲ���������֬��ϳɣ� �����쳣֬����(ʮ��̼��ʮ��̼��ʮ��̼֬����)���������������䡢����(�ٴ��ϳ�Ϊ�Ǽ��Ժϲ�����֢)��
2.ω-������֬�����ω-�������ڸ������н��У��ɼӵ���ø���ġ�������֬�����ω��̼ԭ���ǻ�����ω-��֬���ᣬ�پ�ωȩ֬��������α��ω-�����ᣬȻ����α-�˻�ω-�˻���������������β-�������������������CoA��
����3.α-������֬�������������ɼӵ���ø������ø������α-��֬�������һ��̼ԭ�ӵ�֬����Ĺ��̳�Ϊ֬ �����α-����������֬�����ɼӵ���ø�����ɿ���Ѫ�������Ҷ������������O2��Fe2+����������α-��֬���ᣬ��������֬����֬����Ҫ�ɷ֣�α-�� ֬��������������Ⱦ���������̼ԭ��֬���ᡣα-�����ϰ��߲�������ֲ����(phytanic acid,3��7��11��15-�ļ�ʮ������)��ţ�̺Ͷ���֬���о��д˳ɷ֣��������ڴ����ѻ�������Refsum�ϲ���α-������Ҫ������֯�ڷ����� ���α-�����ϰ���������֢״��
����4.������֬����(unsaturated fatty acid)��������������Լ��1/2���ϵ�֬�����Dz�����֬���ᣬʳ����Ҳ���в�����֬���ᡣ��Щ������֬�����˫������˳ʽ�ģ����ǻ�����β-���� ʱ������3��˳ϩ֬��CoA����ʱ��Ҫ˳��3����2�칹ø��ʹ������2����ϩ֬��CoA�Ա��һ����Ӧ��2����ϩ֬��CoA��ˮ������D��β-��֬�� CoA����Ҫβ-��֬��CoA�����칹ø����ʹ����D������ת���L�����ͣ��Ա��ٽ���������Ӧ(ֻ��L��β-��֬��CoA������Ϊβ-��֬��CoA�� ��ø�ĵ���)��
����������֬������ȫ��������CO2��H2Oʱ�ṩ��ATP������̼ͬԭ�����ı���֬���ᡣ
����
����ȴ�������֬���� ��л 37��ҽѧ��
��һƪ����֬��л
��һƪ������������л��37��ҽѧ����










