红细胞的代谢
2012-05-16 20:23:45 来源:37度医学网 作者: 评论:0 点击:
一、血红素的生物合成
成熟红细胞中,血红蛋白(hemoglolin,Hb)占红细胞内蛋白质总量的95%,它是血液运输O2的最 重要物质,和CO2的送输亦有一定关系。血红蛋白是由4个亚基组成的四聚体,每一亚基由一分子珠蛋白(globin)与一分子血红素(heme)缔合而 成。由于珠蛋白的生物合成与一般蛋白质相同,因此本节重点介绍血红素的生物合成。
血红素也是其它一些蛋白质,如肌红蛋白(myoglobin),过氧化氢酶(catalase),过氧化物酶 (peroxidase)等的辅基。因而,一般细胞均可合成血红素,且合成通路相同。在人红细胞中,血红素的合成从早动红细胞开始,直到网织红细胞阶段仍 可合成。而成熟红细胞不再有血红素的合成。
(一)血红素的合成通路(过程)
血红素合成的基本原料是甘氨酸、琥珀酰辅酶A及Fe++。合成的起始和终末过程均在线粒体,而中间阶段在胞液中进行。合成过程分为如下四个步骤:
1.δ-氨基-γ-酮戊酸(δ-aminplevulinic acid,ALA)的生成:在线粒体中,首先由甘氨酸和琥珀酰辅酶A在ALA合成酶(ALa synthetase)的催化下缩合生成ALA。ALA合成酶由两个亚基组成,每个亚基分子量为60,000。其辅酶为磷酸吡哆醛。此酶为血红素合成的限 速酶,受血红素的反馈抑制。
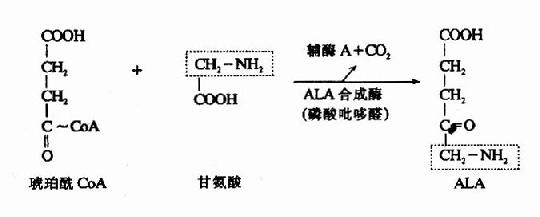
2.卟胆原的生成:线粒体生成的ALA进入胞液中,在ALA脱水酶(ALa dehydrase)的催化下,二分子ALA脱水缩合成一分子卟胆原(prophobilinogen,PBG)。ALA脱水酶由八个亚基组成,分子量为26万。为含巯基酶。
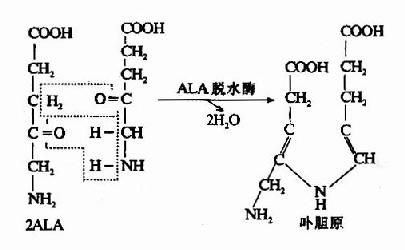
3.尿卟啉原和粪卟啉原的生成:在胞液中,四分子PBG脱氨缩合生成一分子尿卟啉原Ⅲ (uroporphyrinogen Ⅲ,UPG Ⅲ)。此反应过程需两种酶即尿卟啉原合酶(uroporphyrinogen synthetase)又称卟胆原脱氨酶(PBG deaminase)和尿卟啉原Ⅲ同合酶(uroporphyrinogen Ⅲ cosynthase)。首先,PBG在尿卟啉原合酶作用下,脱氨缩合生成线状四吡咯。再由尿卟啉原Ⅲ同合酶催化,环化生成尿卟啉原Ⅲ。无尿卟啉原Ⅲ同合 酶时,线状四吡咯可自然环化成尿卟啉原Ⅰ(UPG-Ⅰ),两种尿卟啉原的区别在于:UPGⅠ第7位结合的是乙酸基,第8位为丙酸基;而UPg Ⅲ则与之相反,第7位是丙酸基,第8位是乙酸基。正常情况下UPG-Ⅲ与UPG-Ⅰ为10000:1。
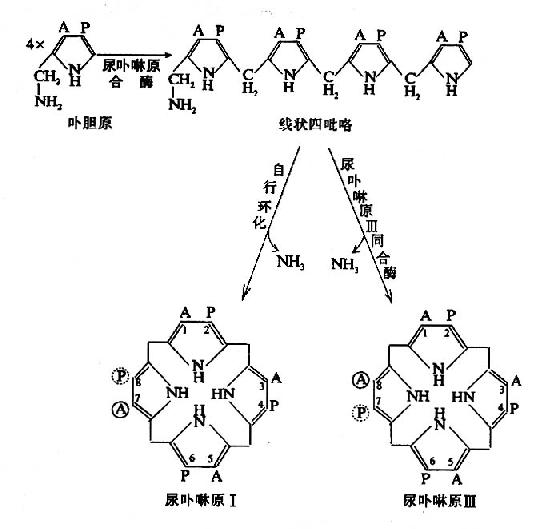
式中A代表乙酸基,P代表丙酸基
尿卟啉原Ⅲ进一步经尿卟啉原Ⅲ脱羧酶催化,使其四个乙酸基(A)脱羧变为甲基(M),从而生成粪卟啉原Ⅲ(coproporphyrinogen Ⅲ,CPG Ⅲ)。
4.血红素的生成:胞液中生成的粪卟啉原Ⅲ再进入线粒体中,在粪卟啉原氧化脱羧酶作用下,使2、4位的丙酸基 (P)脱羧脱氢生成乙烯基(V),生成原卟啉原IX。再经原卟啉原IX氧化酶催化脱氢,使连接4个吡咯环的甲烯基氧化成甲炔基,生成原卟啉IX。最后在亚 铁螯合酶(ferrochelatase)催化下和Fe2+结合生成血红素。(图10-18)。

A.乙酸基 P.丙酸基 M.甲基 V.乙烯基
?图10-18 血红素的生物合成及其调节
血红素生成后从线粒体转入胞液,与珠蛋白结合而成为血红蛋白。正常成人每天合成6克Hb,相当于合成210mg血红素。
(二)血红素合成的调节
血红素的合成受多种因素的调节,其中主要是调节ALA的生成。
1.ALA合成酶 血红素合成酶系中,ALA合成酶是限速酶,其量最少。血红素对此酶有反馈抑制作用。目前认 为,血红素在体内可与阻遏蛋白结合,形成有活性的阻遏蛋白,从而抑制ALA合成酶的合成。此外,血红素还具有直接的负反馈调节ALA合成酶活性的作用。实 验表明,血红素浓度为5×10-6M时便可抑制ALA合成酶的合成,浓度为10-5~10-4M时则可抑制酶的活性。正常情况下血红素生成后很快与珠蛋白 结合,但当血红素合成过多时,则过多的血红素被氧化为高铁血红素(hematin),后者是ALA合成酶的强烈抑制剂,而且还能阻遏ALA合成酶的合成。
雄性激素——睾丸酮在肝脏5β-还原酶作用下可生成5β-氢睾丸酮,后者可诱导ALA合成酶的产生,从而促进血红素的生成。某些化合物也可诱导ALA合成酶,如巴比妥、灰黄霉素等药物,能诱导ALA合成酶的合成。
2.ALA脱水酶与亚铁螯合酶:ALA脱水酶和亚铁螯合酶对重金属敏感,如铅中毒可抑制这些酶而使血红素合成减少。
3.造血生长因子:目前已发现多种造血生长因子,如多系(multi)一集落刺激因子,中性粒细胞-巨噬细胞 集落刺激因子(GM-CSF)、白细胞介素3(IL-3),及促红细胞生成素等。其中促红细胞生成素(erythropoiefin,EPO)在红细胞生 长,分化中发挥关键作用。人EPO基因位于7号染色体长臂21区,由4个内含子和5个外显子组成。所编码的多肽由193个氨基酸残基组成。在分泌过程中经 水解去除信号肽,成为166个氨基酸的成熟肽。分子量为18398。EPO为一种糖蛋白,由多肽和糖基两部分组成,总分子量为34000。糖基在Epo合 成后分泌及生物活性方面均有重要作用。成人血清Epo主要由肾脏合成,胎儿和新生儿主要由肝脏合成。当循环血液中红细胞容积减低或机体缺氧时,肾分泌 Epo增加。Epo可促进原始红细胞的增殖和分化、加速有核红细胞的成熟,并促进ALA合成酶生成,从而促进血红素的生成。
此外铁对血红素的合成有促进作用。而血红素又对珠蛋白的合成有促进作用。
血红素合成代谢异常而引起卟啉化合物或其前身体的堆积,称为卟啉症(porphyria)。先天性红细胞生成 性卟啉症(congenital erythropoietic porphyria)是由于先天性缺乏尿卟啉原Ⅲ同合酶,而使线状四吡咯向尿卟啉原Ⅲ的转变受阻,致使尿卟啉原Ⅰ生成增多。病人尿中有大量尿卟啉Ⅰ和粪卟 啉Ⅰ出现。
二、成熟红细胞的代谢特点
成熟红细胞不仅无细胞核,而且也无线粒体、核蛋白体等细胞器,不能进行核酸和蛋白质的生物合成,也不能进行有 氧氧化,不能利用脂肪酸。血糖是其唯一的能源。红细胞摄取葡萄糖属于易化扩散,不依赖胰岛素。成熟红细胞保留的代谢通路主要是葡萄糖的酵解和磷酸戊糖通路 以及2.3一二磷酸甘油酸支路(2,3-biphosphoglycerate,2.3桞PG)。通过这些代谢提供能量和还原力(NADH,NADPH) 以及一些重要的代谢物(2,3桞PG),对维持成熟红细胞在循环中约120的生命过程及正常生理功能均有重要作用。
(一)糖酵解
循环血液中的红细胞每天消耗约30g葡萄糖,其中90~95%经糖酵解被利用。一分子葡萄糖经酵解可产生2分 子ATP。红细胞中生成的ATP主要用于维持红细胞膜上的离子泵(钠泵、钙泵),以保持红细胞的离子平衡;维持细胞膜可塑性;谷胱甘肽合成及核苷酸的补救 合成等。缺乏ATP则红细胞膜内外离子平衡失调,红细胞内Na+进入多于K+排出、Ca++进入增多,红细胞因吸入过多水分而膨大成球状甚至破裂。同时由 于ATP缺乏,可使红细胞膜可塑性下降,硬度增高,易被脾脏破坏,造成溶血。
红细胞无氧酵解中生成的NADH+H+是高铁血红蛋白还原酶的辅助因子,此酶催化高铁血红蛋白还原为有载氧功能的血红蛋白。
(二)2,3-二磷酸甘油酸(2,3-BPG)支路
在糖无氧酵解通路中,1,3-二磷酸甘油酸(1,3-BPG)有15~50%在二磷酸甘油酸变位酶催化下生成2,3-BPG,后者再经2,3-BPG磷酸酶催化生成3?磷酸甘油酸。经此2,3-BPG的侧支循环称2,3-BPG支路(图10-19)。
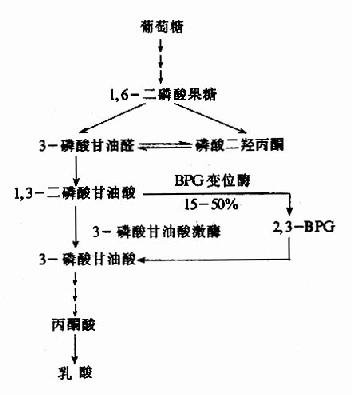
图10-19 2,3-BPG支路
红细胞中2,3-BPG磷酸酶活性远低于BPG变位酶,使2,3-BPG的生成大于分解,因而红细胞中2,3-BPG的浓度处于有机磷酸酯的巅峰,较糖酵解其它中间产物的有机磷酸酯高出数+甚至数百倍(表10-4)。?
表10-4 红细胞中各种糖酵解中间产物的浓度(微克分子/升红细胞)
糖酵解中间产物 动脉血 静脉血 6-磷酸葡萄糖 30.0 24.8 6-磷酸果糖 9.3 3.3 1,6二磷酸果糖 0.8 1.3 磷酸丙糖 4.5 5.0 3-磷酸甘油酸 19.2 16.5 2-磷酸甘油酸 5.0 1.9 磷酸烯醇式丙酮酸 10.8 6.6 丙酮酸 87.5 143.2 2,3-DPG 3400 4940
2,3-BPG能特异地与去氧血红蛋白(deoxy Hb)结合,2,3-BPG进入血红蛋白α2β2四聚体中心空隙两个β亚基之间,借其分子中所带5个负电荷与两个β亚基的带正带氨基酸残基以盐键及氢键结 合,使两个β亚基保持分开的状态,即促使血红蛋白由紧密态向松驰态转换,从而减低血红蛋白对氧的亲和力(图10-20)。
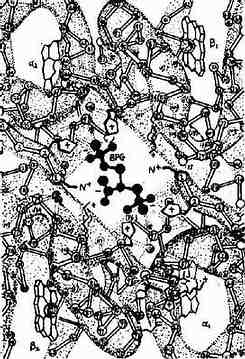
图10-20 BPG与Hb的作用示意图
当红细胞内2,3-BPG浓度升高时有利于HbO2放氧,而2,3-BPG浓度下降则有利于Hb与氧结合。 BPG变位酶及2,3-BPG磷酸酶受pH值调节。在肺泡毛细血管血液pH高,BPG变位酶受抑制而2,3-BPG磷酸酶活性强。使红细胞内2,3- BPG的浓度降低,有利于Hb与O2结合。
反之,在外周组织毛细血管中,血液pH下降,2,3-BPG的浓度升高,则利于HbO2放氧,借此调节氧的运输和利用,具有重要生理意义。但2,3-BPG的生成是以减少一个ATP的生成为代价的。
(三)磷酸戊糖通路
红细胞内利用葡萄糖的5~10%通过磷酸戊糖通路代谢,为红细胞提供另一种还原力(NADPH),NADPH在红细胞氧化还原系统中发挥重要作用,具有保护膜蛋白、血红蛋白及酶蛋白的巯基不被氧化,还原高铁血红蛋白等多种功能。
1.谷胱甘肽代谢,红细胞内谷胱甘肽含量很高(2×10-3mol/L),而且几乎全是还原型GSH)。 GSH的主要生理功能是对抗氧化剂对巯基的氧化。细胞内可自发生成少量超氧阴离子(O-2),同时感染时的白细胞吞噬作用亦可产生O-2,可被超氧化物歧 化酶(superoxide dismufase SOD),催化生成过氧化氢(H2O2)。
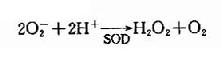
而GSH在谷胱甘肽过氧化酶作用下将H2O2还原为H2O,GSH自身被氧化为氧化型谷胱甘肽(GSSG)。后者在谷胱甘肽还原酶催化下,由NADPH+H+供氢重新还原为GSH。(图10-21)。
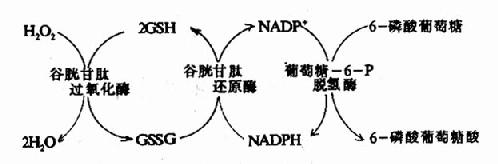
图10-21 谷胱甘肽的氧化与还原
催化NADPH生成的关键酶为葡萄糖-6-磷酸脱氢酶。此酶缺陷的病人一般情况下无症状,但有外界因素(如进食某种蚕豆)影响,即引起溶血。因吃蚕豆可诱导发病,故这种病又称蚕豆病。
2.高铁血红蛋白(methemoglobin MHb)的还原:由于各种氧化作用,红细胞内经常有少量MHb产生,而由于红细胞内有一系列酶促及非酶促的MHb还原系统(表10?),故正常红细胞中MHb只占1-2%。?
表10-5 红细胞中MHb还原系统
还原系统 占总还原能力的百分比(%) 酶促还原系统 NADH脱氢酶Ⅰ 61 NADH脱氢酶Ⅱ 5 NADH脱氢酶 6 非酶促还原系统 抗坏血酸 16 谷胱甘肽 12由表10-5可知,催化MHb还原的主要是NADH-脱氢酶,辅酶为NADH+H+。NADPH脱氢酶,(以 NADPH+H+为辅酶)也参与MHb还作,但作用较小。除此之外,抗坏血酸和GSH可直接还原MHb,而氧化型抗坏血酸和GSSG的还原作用最终需 NADPH+H+供氢。








-脂溶性维生素")

论坛新帖
医学推广
频道本月排行
热门购物
评论排行
- 2011年临床执业医师考试实践技能真...(13)
- 腋臭手术视频(11)
- 2008年考研英语真题及参考答案(5)
- 节食挑食最伤女人的免疫系统(5)
- 核辐射的定义和单位(5)
- CKD患者Tm与IMT相关(5)
- 齐鲁医院普外科开展“喉返神经监护...(5)
- windows7激活工具WIN7 Activation v1.7(5)
- 正常微循环(5)
- 美大学性教育课来真的 男女上阵亲...(4)
