�����֯����
2012-04-12 13:32:57 ��Դ��37��ҽѧ�� ���ߣ� ���ۣ�0 �����
�����֯�������зֲ���Ϊ�㷺��һ����֯�������ǡ��������ǡ����졢�ʹ���Ƥ�����ʼ�Ѫ�ܵ�(��13��1)��������ص��� ϸ���ٶ����ʶ࣬��ϸ������һ���ɻ���(ground substance)����ά(fiber)��������ɡ�����Ϊ���εĽ�̬���ʣ���Ҫ�ɷ�Ϊ������(Proteoglycan)����ά������ԭ��ά���� ����ά����״��ά���ֱ��ɽ�ԭ���ס����Ե�����״�����ɡ��������ؽ��ܵ����Ǻͽ�ԭ���ṹ�����ܼ���л����
��13��1�����ֽ����֯���������ܺʹ���ӽṹ
(ȡ��Karlson��Gerok��Groβ������������ѧ��)
��֯���� ��е���� ������ ̼ˮ����* ���� ��������** ���ͽ�ԭ���� ���������� ����ѹ��ά�����Σ� ������ ��������� ������ ��ѹ������Ħ�� ���ͽ�ԭ���� ���������� ���Ժ�����������أ� ���� ����ǿ�ȴ� ���ͽ�ԭ���� ����Ƥ���� ���ԣ����ԣ�С
���������� ��Ѫ��
����ǿ ���Ե��� ���������� ������ǿ ���ͺ͢��ͽ�ԭ���� �����ᡢ����Ƥ���ز���ʶ������������ ������***�� �ؽ�Һ ���� ���ͽ�ԭ���� ������ Ƥ�� ���ж����Ժͱ����Զ� ���ͣ�80��������ͽ�ԭ���ס� ����Ƥ���� ������ �ǵ��� ������ ����Ĥ ���������á��ָ����á� ���ͺ͢��͵���ԭ���ס����� �����������أ����� ѡ������ ����ճ������ ��Ĥ ������� ��������ͽ�ԭ���� ��������� �����أ����������أ�
����*��Ҫ�ɷ�(����50%)�ú�����ӡ��������10%������������С�
����**�Ը��γ���(����ʯ)�ơ�
����***�ʴ�ϸ���ĵ��Ͷ��ǡ���
��һ�ڡ�������
���������֯�����е�����������Թ��ۺͷǹ��ۼ��������ɶ��־���ӳ�Ϊ������(proteoglycans)��ճ����(mucoproteins)�����������Զ�����Ϊ���������ʲ�����ռ������С������һ��������������������������������ɴ����������ϡ�
����һ����ѧ�ṹ
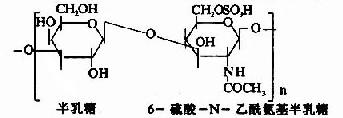
�����������еĶ�����Ϊ�Ӷ��ǣ�������ɳɷ��о����������ǣ����Գ�Ϊ�������ǻ��ǰ����� (glycosaminoglycans)�����������֯�г����İ������ǰ���������(hyaluronic acid)������������(chondroitin sulfate)�����������(keratan sulfate)����(heparin)�ȡ�
��13��2�������г������ǰ�����

���� �����ṹ��λ ��Ҫ���ڲ�λ ������ �ؽ�Һ�����ǡ������֯���ʡ�Ƥ���������������Һ 4-���������أ�����������A�� 
�ǡ����ǡ���Ĥ��Ƥ����Ѫ�� 6-���������أ�����������C�� 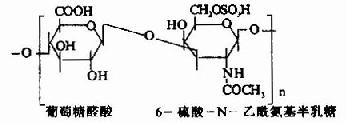
���ǡ����졢��������� ����Ƥ���أ�����������B�� 
Ƥ�����ʹ����������İ�Ĥ ��������� ��Ĥ�����ǡ���� ����

������������������B�ֳ�Ϊ����Ƥ����(dermatan sulfate)�������ǰ����ǵĽṹ��ԪΪ���ǵ�λ�������������������Ǻ���ȩ��(������������⣬������ȩ�����֮������)�����������ⶼ���� �ᡣ����ǰ�����Ϊ���ԣ���֮���Ӵ����ճ�ԣ����ֳ�Ϊ����ճ����(acidmucopolysaccharide,AMPS)��
��������������ͼ�μ�ʵ���Ʋ⣬������Ϊ“ƿˢ״”���ӽṹ(ͼ13��1)���䵰�����ǵ�λ(“ˢë”)�ǹ��۸�����������������(backbone)��������200��300A��
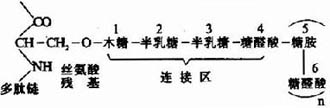
ͼ13��1�������ǵķ��ӽṹ
�����������ǵ�λ��һ�����ĵ���(core protein)�����������ϵ��ǰ�������ɣ�������ҪΪ��������غ����������ء��������ж��ֲ�ͬ�ĺ��ĵ��ף���������200��300kD���������� ֯ϸ���з��ڵ�����һ�ֶ��ġ����ĵ��߶���չN-ĩ�ˣ��γ�һ��״����Լ60-70kD���ǹ������������������ϣ���һ��Լ40-60kD�����ӵ� ��(link protein)�����ȶ���״�������������ķǹ������ӡ����ĵ���������ɵ��ǵ�λ�ɷ�Ϊ������
����(1)N��������������״�����������н��ٵĹ�������
����(2)������������Ϊ��������ع���������Ҫ�����������������������ں��ĵ�������˿������հ���л�������ԭ���ϡ�
����(3)C��ĩ�������������������ء�ͨ�������Ǫ������Ǫ�ľ�����������ں��ĵ���˿����л���
���������ǵ���������������Ϊ4000��40��000�����ɸ����ϰٸ����ĵ��ף�ÿ�����ĵ��ɽ��50���������������100�����������������ɴ˿ɼ������Ƿ��Ӿ������ɸߴ���ǧ��
�������������ǵ���������
���������Ƿ��Ӵ߸߶���ˮ�ԣ��Ա��ֽ����֯ˮ�ּ�����֯�����ʽ���������Ҫ���á�����������֯�н�ԭ��ά ���г�����״�������϶����䵰���ǣ������и߶���ˮ�ԣ���������ˮ�������У���������ѹʱ��ˮ�ֿɱ���ѹ��ȥ������ѹ���ֿ������������ؽ�������Ѫ�� ��Ӧ����Ӫ�����ʵĽ�����Ҫ���˶�����ѹ���仯ʹҺ���������ɴ˿��Խ��ͳ��ڲ��˶����¹ؽ����ǵ�ή����
���������ǵ������Ϻ��н϶�����Ի��ţ�����ϸ����Һ��Ca������Mg������K����Na�����������нϴ������������ܵ�����Щ����������֯�еķֲ���
���������Ƿ��Ӿ��нϴ��ճ���ԣ���������֯���棬�ܻ�����֮֯��Ļ�еĥ��������������������á�����ؽ�Һ�еĵ��������ؽ�������á����ʪ�ؽ������ߣ��ؽ�Һ�е����Ǽ��٣�ʧȥԭ�������ã�����ؽ����ױ��ƻ���
�����������봴�˵������������й�ϵ��Ƥ�����˺����ѿ�γɹ����У�ͨ�������ǰ����ǵ������������ٽ���ԭ��ά �ĺϳɣ�������в���������������ֵ����ǿɵ���ijЩ�����������ӵ����á������������άϸ����������(basic fibrotlast growth factor,bFGF)����ڵ����ǵĸ��ػ�������������ϣ�����Ҫ��bFGF���ǰ����Ǹ�������ʽ��ϸ������������ϡ�bFGF����ػ���������� ����ܱ���bFGF�ⱻ���⡣���˵���ˮ�����ػ���������������ֽ���ʱ���ͷų�����bFGF���ǰ����Ǹ�����̼�ϸ����ֳ���ֻ�������һЩ�������� Ҳ���������û��ƣ������ԣ��㷺�ʹ������ڵĵ����ǿɾ�����Щ���������������ϸ���̾��������á��������֯�����нṹ������Ҫ���塣
�������������ǵ�����ϳ�
���������ǵĺϳ����Ȱ�����������ϳɵ�ԭ�����ں������Ϻϳɶ��ģ����������������У��������������ι����У� ����Ӧ��ת��ø�����Ե���ת�Ƶ�������IJ����ϣ��ϳɰ������ǡ�������������ͼӹ������ڸ߶�������С�����˵���ĵĺϳ���רһ������ƣ����������ǵ� �ϳ���Ҫ��ø�ķָ���λ�Ͷ�ø��������������
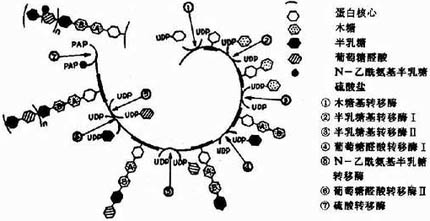
�������밷�����Ǻϳɵĸ��ֵ��Ǽ������������Ȼ�ɻ��Ե��ǣ��������������(UDP)��ϣ������ֵ��Ǽ��������������������ת�����������ͼ13-2��ʾ��

ͼ13��2�����ֵ��Ǽ������������Դ
�����ǰ����Ǻϳɵ���ʼ��������ľ��ת��ø(Xylose transfevase)�Ĵ��£���һ����ľ�ǻ����ӵ����ĵ���������˿����л��ϣ��γ�O�����ռ�(O��glycosidic bond)�����ɰ�����ת��ø(galactose transforase)������ת�������Ӱ����ǣ�����“ľ��뗰�”������������Ȼ�����ɸ߶�������ǻ�ת��ø������˳���ӳ��������ϳɺ��ٽ�һ�� ���Ρ��ɲ����칹ø����������ȩ��ת��Ϊ������ȩ�ᣬ������������ɻ��������(PAPS)�ṩ��������ת��ø
ͼ13��3���ǰ��������������������
�������ӵ��ºϳɵĶ������İ������ǻ��ϡ�(ͼ13-4Ϊ����������A�ϳɹ���ʾ��ͼ)����Vit Aȱ��ʱ����ת��ø�����½����ǰ����Ǻϳ����ޣ���������֯�������������ϰ���
ͼ13��4������������A�ĺϳ�ʾ��
�����ġ������ǵķֽ��л
���������֯�����еĵ�������Ҫ����֯����øD�ȵ����ã���������ˮ������Ĵ���������СƬ�οɱ�ϸ�����ɣ���������ø������ˮ��ɸ��ֵ��Ǽ����������ˣ���ø�����ǰ����Ƿֽ����Ҫ������

ͼ13��5���������ˮ��
������ø���зֽ��ǰ����ǵ�ø������������ø����������ø��������ø�ȡ���ͼ13-5��ʾ�������ˮ����̣��� ��������ø(hyaluronidase)Ϊһ������ø����ˮ�������ᡢ����������A��C�е�β-N-���������������ռ���������ҪΪ���ǻ����ǵĹ� �ǡ��������β-������ȩ��ø��β-N-����������������ø������ø��һ��ˮ�⣬��Ϊ���Ǽ��������
����������ǰ����Ƕ�����������š���ø����Ҳ���ڶ���������ø����ˮ�����������������Щ����ø�к�ǿ�ĵ��������ԡ�����ˮ���������Ĺ���������ˣ�������ø��������ȱ���������ǰ����Ƿֽ��ϰ�������ճ����֢(mucoplysaccharidoses)��










