�Դ�л��ijЩ�ص�
2012-02-29 21:32:04 ��Դ��37��ҽѧ�� ���ߣ� ���ۣ�0 �����
һ��������Ӧ(Energy Supply)
�����ԵĻ˲Ϣ��䣬��Ҫ���������ļ�ʱ��Ӧ����ϸ������������߷���(���ἰ������)�ĺϳ��Լ����ʵĺ� �����ͷŹ�Ȼ���Ǻ��ܵĹ��̣�����Щ�в�����˵��Ϊʲô��ϸ���Ĺ��ܻ��֮������֯ϸ��Ҫ���ĸ�����������Ե�����������Ҫ���ھ������ϵذ�Na���ó� ϸ���⣬ʹȥ����(depolarization)���ĤѸ�ٻָ�Ĥ��λ����ά�����˷ܺʹ������ԵĴ�л��(metabolic rate)�Ǻܸߵģ��������õ�λʱ��ĺ�����(oxygen consumption)�ͻ����������������������ָ������ʾ�����14��1��
��14-1�������������ӵ���Ѫ�����ʹ�л��
�� ���� �� ÿ�ٿ�����֯ ȫ�� Ѫ������ml/�֣� 57 798 ��������ml/�֣� 3.5 49 ���������ģ�mg/�֣� 5.5 77 CO2���ɣ�ml/�֣� 3.5 49������Ѫ����ռ���������15%��������ռȫ���ܺ�����(Լ250ml/��)��20%��Ȼ���Ե�����ֻ����ռ���� ��2%�������������������֯ѪҺ�Ļ�ѧ�ɷ�(���������)���֣������������⣬��������Ϊ��Դ������û�����Եļ��١����ϱ��ɼ�ÿ�ٿ�����֯ÿ���Ӳ� ���Ķ�����̼�����ĵ�����Ϊ3.5ml(156�˷���)��������(respiratory quotient)Ϊ1�����ߣ�����ѧ����(1�˷�����������ȫ����Ҫ����6�˷��ӵ���)�������ǵ������ʻ��Ը������������ʡ�
������Щ��ʵ�ۺ�����˵��������֯���������ǵ����������ܵģ���������˵�����������������£�����֯Ψһ�������� Ϊ��Դ����Ϊ������ԭ��������(С��0.1%)�����Ա�������Ѫ�ǵĹ�Ӧ����Ȼ����֯����������ͪ�壬�������Ե�Ѫ��Ϊǰ�ᣬ�����ڼ�������ͪѪ֢ (ketonemia)������¡����Ѫ�Ǻ�Ѫͪ�������ʱ(����ͪѪ֢)������Ȼ���������������Թ��ܡ�������Ϊ��������ͪ������Դ�ǶԼ���ʱ��Ѫ�ǵ���Ӧ���������Լ����IJ��ˣ��Եĺ�������һ�����������ͪ�塣
������ϸ�������������ǽͽ�(EMP)øϵ�����Ǽ�ø����ԼΪ������֯��20�������Ǽ�ʹ���̶ȵط����ǽͽ�� ����Ҳ�������㹩�ܵ���Ҫ�������������ǵ������������������Ĺ���һ��Ҳ�����жϡ���������֯��Ҫ�����ǵ���������������������������ȱ�Ǻ�ȱ�������� �С�Ѫ���½�50%�����»��ԣ����ж�(�����Ե�)Ѫ�������ӾͿ������������ٴ���ʹ�ô�����ȵ�����Ϊ�����ֶ�Ҳ����������ԡ��ȵ��ض��Ե�����Ӱ�죬 ������Ϊ�Ǽ�����õĽ����Ҳ������Ϊ�ȵ��ؽ�����Ѫ��֮�ʡ���Ϊ�ȵ����Ǵ�������ʣ���������Ѫ�����ϣ���Ͷ�������֯����Χ����֯�����ò�ͬ���� ��Щ��֯���ȵ��ص����ÿ�����Ҫ��ֱ��Ӱ����������ǵ���ȡ��
��������ATP��ˮƽ���ߣ����ĺϳɺ����þ���Ѹ�١��ݲⶨ������ATPĩ��������İ�������ʱ��ƽ��ֻ��Լ3�� �ӣ�����֯�����ἡ��(CP)ˮƽ��ATP��Ҫ�ߣ����ɿ�����ATPĩ�˸����������һ��������ʽ�������ἡ�ἤø(CPK)�Ĵ��£�ATP��CP���� ��ת�䡣

��������һ�����ܷ�Ӧ��������ת���в������������Ķ�ʧ������CPK���Խ�ǿ������������������Ͷ��á����⣬����֯�л��л��Խ�ǿ�ļ���ø(myokinase)�����ɴ����з�Ӧ���˷�Ӧ����������“�ھ�DZ��”�����������ATP�е����������������
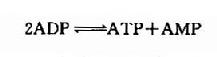
�����������ּ�ø����ά�����ڸ�ˮƽATP��������֤��
����������֬����ɺʹ�л
������֬����֯�⣬����ȫ����֬��������֯����֬����֯��Ҫ����������(����֬)��������֯�е�֬�༸��ȫ���� ֬���Ը��ص�1��2��֬�࣬���Ǿ�ȫ��ƽ�����ԣ�����ֱ�ⶨ�Ի��ʺ����ʵĻ�ѧ�ɷ֣��ͻᷢ�ֻ��ʺ�ˮ�ݺ͵����ʽ϶࣬֬���ռ���ص�1��3�������� �е�֬�ຬ���϶࣬Լռ���ص�55%��
��14��2�����������Ե�֬�����
�ɷ� ���� ���� ���أ����� ���أ����� ��֬������ ���أ����� ���أ����� ��֬������ ˮ 81.9 71.0 ��֬ 5.9 32.7 100 15.6 54.9 100 ����֬ 4.1 22.7 69.5 7.2 25.2 45.9 ���̴� 1.3 7.2 22.0 4.3 15.1 27.5 �ܰ�������֬ 0.4 2.4 7.3 4.1 14.5 26.4 �������� 0.3 1.7 0.05 0.18�������ϱ��ɼ�����ʪ�ؼƣ�������֬�ຬ��Լ�����ڻ��ʡ����ֲ����Ҫ�����ڰ����е�����ά�ⱻ������(myelin sheath)�������ʵ�֬��ɸߴ���ص�70-80%��
�������е���֬��Ҫ���Թ�����Ԫ(neurone)����Ĥ�����ʡ���ЩĤ�Խṹ��������֯ϸ����Ĥ�ṹ�й�֮ͬ ��������������֬�뵰���ʹ��ɵĸ����������֬����ɺʹ�л������һЩ�ص㡣���������ʣ�������ijЩ�������֬�ɷ֣���Щ�ɷֻ��߽��������ʣ��������� �к����϶࣬����������֯������ټ���������ȩ��֬(plasmalogen)������֬(cerebroside)��
�����Բ���Ѫ����ȡ֬���ᣬ����Ҳ��������֬���ᣬ��ô�����Щ��֬��֬�����Ǵ�����������?ʵ��֤�������е�֬����͵��̴�����������CoA�ϳɣ�������CoA����Ҫ��Դ���������ǡ�
���������γ�(Myelination)֮ǰ��δ���������֯�����̴�����֬�϶࣬��������֬���٣�����֬�ϳ�ø ϵ�Ļ���Ҳ���͡��������γ�ʱ����øϵ�Ļ������ߣ�����֬�ĺ�������Ӧ���࣬�����γ�����ϵͳ�ķ�������������أ����������� (demyelination)����ϵͳ��������Ҫ�IJ����ı�֮һ�����ʵĴ�л�ص������ڽ��������γ�ʱ��л�ܿ죬һ���γ�֮��ͱ�ú�������Ϊ������ �ȶ���һ�ֽṹ������Ϊ��������������ȱ������֬�ֽ��л��øϵ���Ѿ��γɵ����ʣ����˸���ɷ�(�������ἡ����֬)�нϸߵĸ������⣬������֬�͵��� ���ȵĸ����ʾ����͡�
���������Ȱ���Ĵ�л�빦��
�����Ե����백���������Ѫ���к����ԵIJ����������Ѫ�����ϵ��ص���Ա����������л�ص���ɵġ��Ժ�Ѫ����ijЩ���백����ĺ����Ա����14-3����
��14��3�����Ժ�Ѫ����ijЩ���백����ĺ���
������ �� Ѫ�� �˷��ӣ��� �˷��ӣ����� �Ȱ��� 10.6 0.05 N-�������Ŷ����� 5.7 - �Ȱ����� 4.3 0.7 γ���������� 2.3 - ���Ŷ����� 2.2 0.01 ɫ���� 0.05 0.05�����ɱ�14-3�ɼ����������백�����ԹȰ���(Glu)������ߣ���������Ѫ���е�Ũ��Ҫ�߳�200�����ϡ��Ȱ��ᡢ�Ȱ�����(Gln)��γ����������(GABA)���ߺ����ܺ�Լռ�������백����������һ�롣���ԣ����Եİ������л�У��Ȱ���ռ����Ҫλ�á�
����Ȼ�����Ȱ�������ͨ��Ѫ�����ϣ����ڹȰ�����Դ�������ĺϳɣ�ͬλ��ʾ��ʵ��������ڹȰ���ϳɵ�ԭ�������� �ǣ�������Ѫ�ǡ������ǽ�����ϸ������ת���α��ͪ�����(α��KG)�����߿��ڹȰ�������ø�Ĵ���ת��ɹȰ��ᣬ��ɾ�ת�����������ɹȰ��ᣬһ���� Ϊ��һ;�����к�ʵ�ʣ���Ϊ�Ȱ�������ø(GDH)���淴ӦʱK��mNH4��Ϊ8mM��Զ����ϸ���ڰ�֮Ũ�ȡ��Ȱ����ڹȰ������ϳ�ø���������백��� ��Ϊ�Ȱ�����������һ�����ܷ�Ӧ(����ATP)�����йȰ������ϳ�ø�Ļ���ǿ����K��mNH4����Ϊ0.39mM�������ɵĹȰ���������Ȱ��ͬ������ ͨ��Ѫ�����϶�����Ѫ�У�����������֯��Ѫ�����������ǣ�ͨ����л����ѪҺ�ԹȰ���������������еİ������ⰱ�Ļ���Σ���ԵĹ��ܡ�

ͼ14��1�����йȰ����л�백������
�������йȰ�
���л����һ���ص�����������γ����������(����γ��������GABA)�����˷�Ӧ��ø�ǹȰ�������ø(GAD)������Ҫ��������ȩ����ø��GABA��һ�������Ե����ʣ�������������ϵͳ������GABA��Ҫ���ڻ��ʣ��ر�����״�塢���ʡ�С�Եij�״�˵ȴ���
����GABA��������Ԫ���ձ����������á�1963�������������GABA��������ͻ��ǰ��ĩ�ң������˷��� ���ʵ��ͷţ��Ӷ��������ơ�����ЧӦ��Ϊͻ��ǰ����(presynaptic inhibition)��GABA�ڼ����е����þ�����ͻ��ǰ����Ϊ������������GABA��Ҫ������ͻ��������(postsynaptic inhibition)��˯��ʱƤ���ͷ�GABA���࣬���������ΪGABA������˯�ߡ����ѵ����������йء�
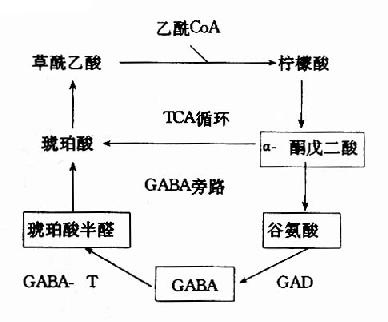
ͼ14��2a������TCAѭ����GABA��л��·
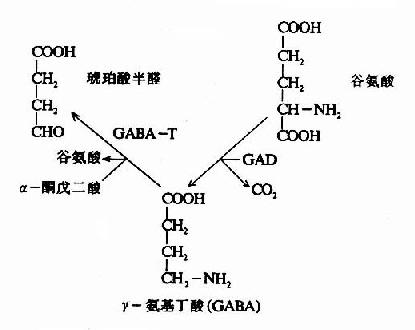
ͼ14��2b��GAD��GABA��T������
��������Ԫ�����ͻ��(synapse)���������ں��д�����γ����������ת��ø(GABA��T)���� �ɴ�GABA��α��ͪ�����֮���ת�����ã������������ȩ(succinic acid semialdehyde)�Ȱ��ᡣ��ɿ�����GABA����һ�ַ�ʽ��GABA��TҲ����Ҫ��������ȩ����ø������GAD�Ƚϣ���ͬ��������ȩ���� �������Ե�����ά����B6ȱ��ʱ����ҪӰ��GAD�Ļ��ԡ����磬ʹ�����������ƽ�˲�ʱ����������������ά����B6(����ȩ)��ϳ������� (isoniazone)������ά����B6��������й����������֯��ά����B6Ũ���½���GAD�������½������GABA�ĺϳ����裬����ʹ��������˷� �������鴤��֢״�����Գ���ʹ��������ʱӦ�ϲ�ʹ��ά����B6�����⣬�ٴ��϶��ھ��ʡ�����Ż�µIJ��ˣ�Ҳ��ʹ��ά����B6�������Ҳ���������֯�� GAD�Ļ��ԣ�ʹGABA�������࣬����������Լ�ǿ��
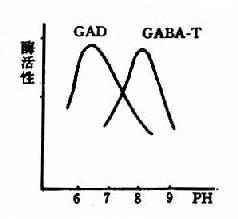
ͼ14��3��GAD��GABA��T����������pH
����GABA��ת�����ú�IJ����������ȩ���������������ᣬ���߽���������ѭ�������������á���ˣ�������֯�е�������ѭ������ϵ��������һ��GABA��л��·(GABa shunt)��
�����Ȱ�������ø��γ����������ת��ø��Эͬ���öԱ�������GABAһ��Ũ������Ҫ���塣����ø������pH�� ͬ��GAD������pHΪ6.5����GABA��T��ΪpH8.2��(ͼ14��3)�ɴ˿ɼ�����ϸ����pH���б䶯�Ϳ����Ըı�������ø�Ļ��ԶԱȡ������ж� ʱ������GAD������ǿ��GABA��T���Լ�������������GABAˮƽ�����������������ƣ���֮�������ж�ʱ����GABA��T������ǿ��GAD���Լ����� ����GABAˮƽ�½������ڷ������Ρ�
��������ָ�����Ȱ�������������˷����ã��������Ȳ���GABAȴ���������ã����ԹȰ���Ĵ�л��������˷ܺ����Ƶ����йء����⣬ͨ��GABA��л��·��Ҳ���Ե�������л���˷����ƹ�����ϵ�����ˡ�
����ȴ��������Դ�л
��һƪ������(Neurotransmitter)�Ĵ�л
��һƪ������֯����










