����(Neurotransmitter)�Ĵ�л
2012-02-12 17:11:03 ��Դ��37��ҽѧ�� ���ߣ� ���ۣ�0 �����
��ϵͳ�ڴ��������ѧ���ʣ������Dz�һ�����ǵ���(transmitter)����Ϊ���ʣ�����߱��������������� (1)����Ϊ�ǵ��ʵ�����Ӧ�����Եش������Ը�����Ϊ���ʵ���Ԫ�У����ң���������Ԫ��ĩ���кϳɸõ��ʵ�øϵͳ��(2)��������ĩ���ںϳ��Ժ� ͨ���Ǽ�������������(vesicle)�ڣ��������Է�ֹ�������ڵ�����ø���ƻ���(3)���嶯����ʱ����ĩ���ڵĵ��ʾ���ͻ��ǰĤ�ͷ���ͻ���� ϶��(4)����ͨ��ͻ����϶��������ͻ����Ĥ�Ľ�����������ⲿλ������ͻ����Ĥ����ͨ�Ըı��Լ���λ�仯��(5)�����ڷ�������ЧӦ��������Ӧ ��Ѹ����ֹ���Ա�֤ͻ�����ݵĸ߶������õ���ֹ�м��ַ�ʽ��һ�DZ�ø��ˮ�⣬ʧȥ���ԣ�����DZ�ͻ��ǰĤ“����ȡ”������һ����Ϊ��Ĥ����ȡ��Ҳ�е� ���ֽ���Ѫѭ������Ѫ��һ���ֱ�ø�������ƻ���
����Ŀǰ��֪����������ܶ࣬����Ҫ���������������Ӱ���(ȥ���������غͶ�Ͱ�)��5����ɫ����GABA��ijЩ��������ĵȡ�
����һ����������(Acetylcholine��дAch)
���������������������������˶���ֲ������ϵͳ�Ľ�ǰ��ά�������ں���ά���˷������ʡ�
����(һ)�ϳɺ�����
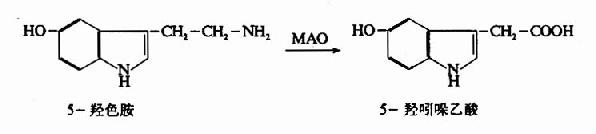
����Ach�ɵ��������CoA���ϳɡ�����������ø(choline acetylase)�����з�Ӧ��
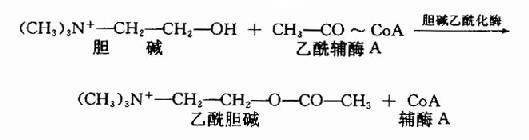
�������ڵ���������øλ�ڰ����ڣ��������Ach�����ڰ����ںϳɣ�Ȼ������������档ƽʱ�����кͰ����е�Ach��Լ��ռһ�룬�����߿��ܴ���ƽ��״̬�������������Ach��һ�ֽ���͵�(�뵰���ʽ��)�����ͷ�������ʱ�����Ϊ�����͡�
����(��)�ͷš�����ȡ��ʧ��
�������嶯����ͻ����ĩ��ʱ����������ͻ��Ĥ������֮�ںϡ����ѣ���ʱ�����ڽ����Achת��Ϊ������Ach���ͷ���ͻ����϶��ͬʱ����������һ���ְ������ºϳɵ�AchҲ��֮�ͷš�
����Ach������ͻ����Ĥ(ͻ������Ԫ��ЧӦϸ����Ĥ)��������壬��������ЧӦ���Ѿ�ȷ��Ach������һ�ַ�����Ϊ42��000�ĵ����ʣ�ͨ����֬������ʽ������Ĥ�ϡ�
����Ach�ڴ�����Ϣ֮�������ֿ���������ͻ����϶�����м��ٲ�����ͻ��ǰĤ������ϵͳ���������±�����ͻ��ǰ��Ԫ����Ach���ڵ�����ø��������ˮ��ɵ���������ʧȥ���ԣ�Ҳ��һ���־���ɢ���뿪ͻ����϶������������������ĩ���еĴ�л�����ܽ���ͼ(14��4��14��5)��
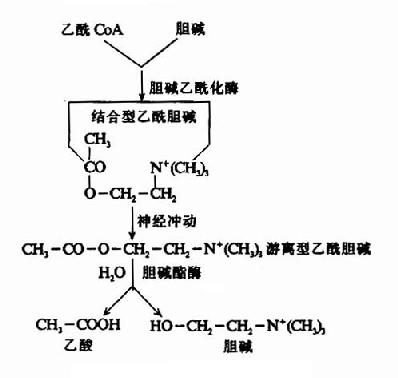

ͼ14��5����������Ĵ�л
ͼ14��4��ͻ����λAch�Ķ�̬
������������Ӱ���(Catecholamines)
��������Ӱ�����ָ�����ڱ����ӻ����ṹ�İ��ࡣ���ھ���������ԵĶ���Ӱ�������Ͱ� (dopamine,DA)��ȥ����������(norepinephrine,noradrenaline,NE)���������� (epinephrine,adrenalin,E)�����ǵĽṹ���¡�

����ȥ���������غ��������ؼ������������������ڵļ��أ����ǽ��к�������ϵͳ��ȥ��������������ά�����ʡ�NE�������ڷֲ��㷺�������϶࣬E���٣�����������ؽ���NE�Ĵ�л��DA����Ҫ����������ϵ��Ҳ��һ�����ʡ�
����(һ)����Ӱ�������ϳ�
��������֯�ж���Ӱ��ĺϳ�ԭ������ѪҺ�е��Ұ��ᣬ��ϳɹ�����ͼ14��6��
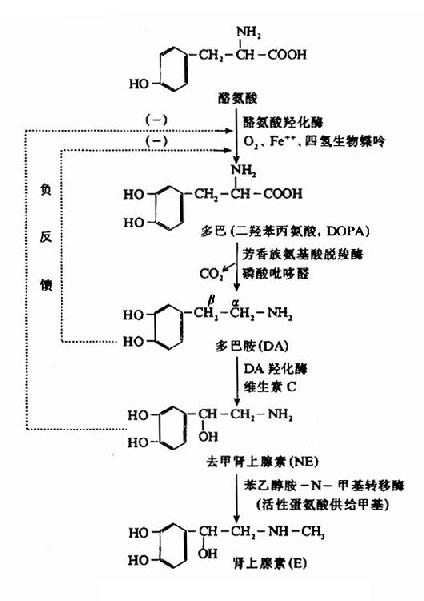
ͼ14��6������Ӱ��������ϳ�
���������������У���һ�����Ұ����ǻ�ø�μӣ���λ��ȥ����������������ά�İ����ڣ������٣����Եͣ���ΪNE ���ɵ�����ø������������������ĸ�ø��O2��Fe����Ҳ�Ǻϳ�ʱ�ز����ٵ����أ��ڶ�����Ӧ���ɰ����з����就��������ø��������ø�������Բ��ߣ��� һ�㰱��������øһ����Ҫ����������ȩ��Ϊ��ø���������Ƕ�Ͱ��ǻ�ø����������Ӧ������������β̼ԭ���ϣ���ø�������ڰ������Ǹ��������ڱڣ����ں� Cu�����ĵ����ʣ�����Ҫά����CΪ�������ӡ�
���������ϸ�ø����ϸ��ˮƽ�ֲ������˽⣬�ϳ�NE�����һ��ֻ���������ڽ��У�NE�ϳ����������Ұ����ǻ�ø���ٵ��ڣ����ҵ���ĩ�Ұ����������NEŨ�ȹ���ʱ�������������������Ұ����ǻ�ø�����ö�����NE�ĺϳɡ�
�������Ҵ�����N����ת��ø��Ҫ��������������ϸ������ʹNE���������������ء���ø�����������������ڣ��� һ����Ϊ����ʱ���������ڵ��������غ������١�������Ϊ��������Ҵ�����N����ת��ø���Թ��ߣ�����ʹ��Ͱ�ֱ�ӱ��N������Ͱ���������Щ���� �Ĵ�л���ң�������Ǿ������֢��ԭ��֮һ��
����(��)ȥ����������(NE)
����1.������ͷš�NE�������ںϳɺ�͵����档����ATP���ȸ������������ɵؽ����һ����������ʹNE ��������������ⵥ������ø���ƻ������嶯����ĩ��ʱ��ͻ��ǰĤ���������ݱ���ǰĤ�ںϣ����Ѳ�����С�ס���ʱ�����ڵ�NE��ͬ�ȸ���������һ�� �ͷ���ͻ����϶��
����2.ͻ����϶��NE��ȥ���ͷ���ͻ����϶��NE����ͻ����Ĥ�ϵ�NE�����ϣ���������ЧӦ��Ȼ��Լ�� 3/4��NE���±�ͻ��ǰĤ����ȡ������������NE�ֿɽ����������档����ȡ��һ�ֺ��ܹ��̣���ͻ��ǰĤ�ϵ�Na����K����ATPøϵͳ������Ĥ�ϵ� Mg������ATPøϵͳ�йء�����һ����NE��Ϊ��Ĥ����ȡ���������ﱻ�ֽ�������һ��������ͻ����϶�б��ƻ�������ѪҺ�����˱�ͻ��ǰĤ���������� ȡ��NE���ٹ������⣬�����NE���ø�ٽ������NE��ø�ٽ�����ͼ14?��ʾ����������ø(MAO)�Ͷ���ӣ�O����ת��ø (catechol��o��transmethylase,COMT)�Ǵ�����Ӱ��ֽ��������Ҫ��ø�����Dz�������������֯�ڣ����ҹ㷺�طֲ��ڷ��� ����֯����Ԫ��������Ĥ��Ҳ�ܶࡣNE��MAO���ã����������Ѱ�������ȩ�������ٱ�ɴ����ᡣ3����������4���ǻ����Ҷ���(MHPG)��������NE ����Ҫ������������������������ݻ�������(VMA)Ϊ����ѪҺѭ���е�NE(��Ҫ�Ǽ���)�����ڸΡ�������֯��COMT���ñ�ɼ�������л������� й�����ڣ��ٴ��ϳ��ⶨ����VMA��������Ϊ�˽⽻�����ܵ�ָ�꣬���ȸ�ϸ��������ĸϸ����ʱ������������֯Ҳ����NE��E�����л����VMA��Ӧ ���࣬����������������塣
��������NE��ͻ����λ�Ķ�̬����ͼ14��8��ʾ��

ͼ14��7��ȥ���������صķֽ��л
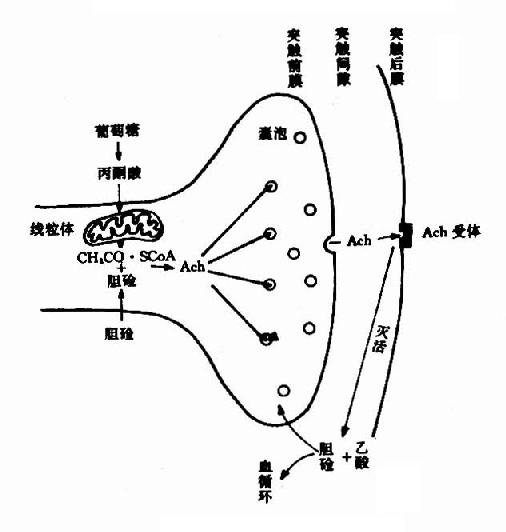
ͼ14��8��ͻ����λNE�Ķ�̬
����������ϵͳ�ڵ�ȥ��������������Ԫ���弯�������Ժ����ԣ���ͨ·�����о���ȷ������NE�����྿�������� �Ի����˷��Եĵ��ʣ�Ŀǰ�в��ܿ϶��������沿λ��ͬ���졣NE�����������ЧӦҲ�����ü�����“�˷�”��“����”���������һЩ����ʵ��۲� ����NE����������˯�����½��ͣ�������ʳ��Ϊ��������Ϊ������NE���٣��ɱ��ֳ�������������֮�������ɱ��ֳ����ꡣ��֮������NE�Ĺ��ܿ��ܺ��� �¡���ʳ��Ϊ����ʹ����Ѫ��ϵͳ�;���״̬�ĵ��������й�ϵ��
����(��)��Ͱ�(DA)
�����ڶ���Ӱ�����ʵ�����ϳɹ����У���Ͱ���ȥ���������ص�ǰ�塣���ڷ���NE����֯�����б�ȻҲ��DA������DA������ijЩ��λ��Ũ�Ⱥܸߣ��������ķֲ�����NE��ƽ�У�����һ����Ϊ������Ҳ��һ�ֶ��������ʡ�
�������ڶ�Ͱ��������Ƕ��ģ������ܺ������˶����ܵļ�ǿ�������ڷ��ڻ��ܵļ�ǿ�Լ������ĵ��ڶ��й�ϵ��
������Ͱ�����ĩ���е�����������DA�ij������������ݲ�ͬ��NE���ݣ�ǰ�߲�����Ͱ���β���ǻ�ø�����Բ��� ��DA�ǻ���NE�����⣬��ȥ��������������ά�У�NE���ݶ��������Ҫ����β��λ�������ǻ�����DA�ṹ��û��β���ǻ���DA�����桢�ͷź�ø�ٽ��ⶼ ��NEʮ�����ƣ��������ٶ����NE�졣����DA�Ĵ�л������Ҫ��3����������4���ǻ�������(homovanillic acid��дHVA������ͬ�������)

��������5����
ɫ����5��ydroxytryptamine ��д5��HT)
����5����ɫ������Ѫ�������(serotonin)�������Ǵ�Ѫ���з��ֵġ�������ϵͳ������5����ɫ������Ԫ�����ڼ������������ϵͳ��������δ������5����ɫ������Ԫ��
��������5����ɫ��������Ѫ�����ϣ����������5����ɫ�������ںϳɵģ������ܵ�5����ɫ������һ����Դ������֯��ѧ�ķ���֤����5����ɫ������Ԫ�İ��������ڵķֲ���Ҫ�����Ըɵ��з��Ⱥ����ĩ����㷺�ֲ����Ժͼ����С�
����(һ)�ϳɡ�������ͷ�
����5����ɫ����ǰ����ɫ���ᡣɫ���ᾭ����ø�ٷ�Ӧ�����ǻ������ȣ�����5����ɫ�����˹�����ij�̶ֳ��ϺͶ���Ӱ����������ơ�

ͼ14��9��5��HT������
����ɫ�����ǻ�ø���Ұ����ǻ�øһ������ҪO2��Fe�����Լ���ø����������ʡ�����������ø�ĺ������٣����Խ� �ͣ���������5��HT����ϳɵ�����ø�����⣬����5��HT��Ũ��Ӱ��ɫ�����ǻ�ø�Ļ��ԣ��Ӷ���5��HT���ŷ��������ҵ������á�Ѫ������ɫ�����Ũ�� ҲӰ������5��HT�ĺϳɣ���Ѫ������ɫ��������ʱ(���������ǻע��ɫ�����)�������Ե�ɫ��������࣬�Ӷ�������5��HT�ĺϳɡ�
����(��)����ȡ�ͽ���
�����Ͷ���Ӱ������һ�����ͷŵ�ͻ����϶��5��HT���ֱ�ͻ��ǰ��ĩ������ȡ����������ȡ���ֽ������������棬������������Ĥ�ϵĵ�������ø(MAO)��������
����
�������������5��HT�������Ҫ��ʽ��5������������(5��hydroxyindolacetic acid)��������ԡ�
�������5��HT�Ը�����Ԫ������ʱ���֣�5��HT��ʹ��������н�ǰ��Ԫ�˷ܣ���ʹ�����н�ǰ��Ԫ���ơ� ��ٶ�����з�˻���ҩ�����5��HT�ϳɣ�����ʹ����5��HT�������Խ��ͣ���������˯���ϰ���ʹ�н��ͣ�ͬʱ����ȵ���ʹ����Ҳ��������ʧ������� �̼�������з�ˣ���Ӱ�����������ߣ���һ���棬Ҳ�۲쵽��������ʱ��������5��HT���¼��١���Щ�����ʾ����5��HT��˯�ߡ���ʹ�����µ��ڶ��й� ϵ�������˱�����5��HT�ܸı䴹����ڷ��ڻ��ܡ����⣬�������5��HT����Ԫ���ƻ��Ǿ����Լ���ʱ���ֻþ���ԭ�ɼ�����Ҳ��5��HT��һ�� �Ĺ�ϵ��
�����ġ����������
�������ڵ����������Ű����ᡣ��ȥֻ��Ϊ�����Ǻϳ���������ԭ�ϣ����ǵ����ʷֽ�IJ����������ע�ijЩ���� ���������ͻ�����������ŵ��ʵ����á����ҷ��֣������������ᣬ��γ���������ᡢ�ʰ��ᡢβ��������ȶ�������Ԫ�����������ã�������������Ȱ� �ᡢ���Ŷ����������Ϊ�˷����á�
������һЩС����������������ϵͳ��Ҳ��������ͬ�����á�1975�귢�ֵ��Է���(enkephalin)�������������л����ɵĹ��ģ��������з���������Է����ַ�Ϊ���͡�
������������(���Է��ģ�Met��enkephalin)��
����H���ң��ʣ��ʣ�����������OH
������������(���Է��ģ�Leu��enkephalin)
����H���ң��ʣ��ʣ�����������OH
��������������ϸ���ںϳɵľ�����������õ��ģ������Է��ġ���֪���(morphine)Ҫ�������ڵ�������� ��ϲ��ܲ�����ʹ���������ã����Է���֮���������������Ҳ����Ϊ����������������ϡ���������������ʣ����Է�������Դ�Եġ����⣬��֪���ڻ���һЩ ������������ϲ�������������õ����������࣬��֮Ϊ�ڷ���(endorphin)����Ȼ������ʮ�ֿ϶���Щ���������������(����ȫ����ǰ���� �����ʵ�����)�����ǣ��о����ǵ����öԲ����ԵĹ��ܣ��ر�Բ���ʹ��ԭ���Ǻ�������ġ�
����ȴ����������� (Neurotransmitter
��һƪ������Ľṹ�빦��The Structure and Function of Nucleic Acid
��һƪ���Դ�л��ijЩ�ص�










