RNA�Ľṹ�빦��
2012-01-26 11:09:00 ��Դ��37��ҽѧ�� ���ߣ� ���ۣ�0 �����
��DNA���Ŵ���Ϣ���������Ŵ���Ϣ������ͨ�����������Ĺ�����ʵ�֣���DNA���ǵ����ʺϳɵ�ֱ��ģ�壬�ϳɵ����ʵ�ģ����RNA������ϸ���Ŵ���Ϣ�������ǣ�
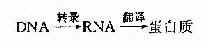
������DNA��ȣ�RNA����࣬��������Խ�С��һ���Ե��������ڣ��������оֲ������ṹ����������ص��� ���������(uridin,U)������������ण������Է�����C��G��U��A֮�䣬RNA������֮����һ���ı�����ϵ����ϡ�м���϶ࡣ�� �⣬tRNA��������ȷ�������ṹ��
��15-7��RNA�ķ���
�� ϸ���˺Ͱ�Һ ������ ���� �˵�����RNA rRNA mt tRNA �˵�������ɳɷ� ��ʹRNA mRNA mt mRNA �����ʺϳ�ģ�� ת��RNA tRNA mt tRNA ת�˰����� ����һ��RNA hnRNA �� ����mRNA��ǰ�� С��RNA snRNA �� ����hnRNA�ļ��ӡ�ת�� С����RNA scRNA/7SL-RNA �� ��������������λ�ϳɵ��ź�ʶ�������ɳɷ�����ע��ԭ��ϸ��������3��RNA��
����(һ)��ʹRNA(mRNA)�벻��һ��RNA(hnRNA)
�����Ŵ���Ϣ��DNA���ӳ�¼��RNA�����еĹ��̳�Ϊת¼(transcription)������������У���� ת¼���ɵ�RNA��Ϊ����һ��RNA(heterogeneous nuclear RNA,hnRNA)��Ȼ����ϸ�����������ã���Ϊ�����ʵİ��������кϳ�ģ�����mRNA(messenger RNA)��hnRNA��mRNA��δ����ǰ�塣����֮��IJ����Ҫ�����㣺һ��hnRNA���������е�һЩƬ�ν�����������Ӧ��mRNA�У���ЩƬ�γ�Ϊ �ں���(intron)������Щ������mRNA�е�Ƭ�γ�Ϊ������(exon)��Ҳ����˵��hnRNA��ת��ΪmRNA�Ĺ����о������ӣ���ȥ����һЩƬ �Σ����µ�Ƭ�α�����������һ�𣻶���mRNA��5′ĩ�˱�����һ��m7pGpppñ�ӣ���mRNA3′ĩ�˶���һ�����������(polyA)β�͡� mRNA��5′ĩ�˵�3′ĩ�˵Ľṹ������5′ñ�ӽṹ��5′ĩ�˷DZ��������������İ��������еı�������3′ĩ�˷DZ��������Ͷ��������β�͡�������� ��βһ������ʮ����һ�ټ�ʮ�����������Ӷ��ɡ�����mRNA����ʱ�����������ξ�Aβ��������̡���ˣ�Ŀǰ��Ϊ����3′ĩ�˽ṹ����������ת¼������ ��ʹmRNA��������ȶ��йء�ԭ�������mRNAû�������ס�β�ṹ��
ͼ15��13��hnRNA��mRNA�Ľṹ�ȽϪ�
(Ϳб����Ϊ�����ӣ��հ���Ϊ�ں���)
����1961�꣬Jacob��Monod���������mRNA�ĸ�������ϸ���У����ڵ��������ڰ����ж������� ���ںϳɣ������ȻҪ����һ���м��ォDNA�ϵ��Ŵ���Ϣ�����������С��������о�֤ʵ�������м��T��ʹRNA����mRNA�ĺ�����������DNA������ Ӧ�������źϳɵ����ʵİ��������С������ָ������������ȷ��˳������������?��ͬ��mRNA�����ɺ�����˳��ͬ������ֻ��A��G��C��U4�ּ� �������һ������Ϳ��Ծ���һ�������ᣬ��ֻ�����ֱ仯��ʽ����������������һ�������ᣬ��ֻ��16�ֱ仯��ʽ������������20�ְ��������Ҫ�� 1961��Crick��Brenner��ʵ��ó����������������һ��������Ľ��ۣ�����������λһ��ĺ������������Ŵ�����(genetic code)�����������룬�����Ϳ�����64�ֲ�ͬ�����룬��������±���ٶ���һЩ������ʹ���������ϵ����롣��һ�ٶ��ܿ�ͱ�֤���ǶԵġ��Ŵ�������� ����������
����(1)�������������һ�������ӣ�ÿ��������������ǰ�������ĺ�������ɣ�һ��������ֻΪһ�ְ�������롣����64�������ӣ�
����(2)������֮�䲻�ص�ʹ�ú����ᣬҲ����������
����(3)һ�ְ�������ж�������ӣ�����ص��Ϊ�����ӵļ��ԣ�
����(4)�����ӵ�ͨ���ԣ������������͵ȵIJ���ֱ�����࣬�����ʺϳɶ�ʹ��ͬһ�������ӱ�(��15-8)�����м��ٵ����⣬������ϸ���������壬Ҷ�������õ��������в�ͬ��(��15-9)����
��15-8��ͨ���Ŵ����뼰��Ӧ�İ�����
��һ��������5′ �ڶ��������� ������������3′ U C A G U �������� ˿���� �Ұ��� ���װ��� U �������� ˿���� �Ұ��� ���װ��� C ������ ˿���� ��ֹ�� ��ֹ�� A ������ ˿���� ��ֹ�� ɫ���� G C ������ ������ �鰱�� ������ U ������ ������ �鰱�� ������ C ������ ������ �Ȱ����� ������ A ������ ������ �Ȱ����� ������ G A �������� �հ��� �춬���� ˿���� U �������� �հ��� �춬���� ˿���� C �������� �հ��� ������ ������ A ������ �հ��� ������ ������ G G �Ӱ��� ������ �춬���� �ʰ��� U �Ӱ��� ������ �춬���� �ʰ��� C �Ӱ��� ������ �Ȱ��� �ʰ��� A �Ӱ��� ������ �Ȱ��� �ʰ��� G��15��9��ͨ���Ŵ��������������Ŵ�����֮���һЩ����
������ ͨ�ñ��� ��������� ���鶯�� ��Ӭ ��ĸ�� ֲ�� UGA ��ֹ�� ɫ���� ɫ���� ɫ���� ��ֹ�� AUA �������� ������ ������ ������ �������� CUA ������ ������ ������ �հ��� ������ AGA ������ ��ֹ�� ˿���� ������ ������ AGA����ע���±������Ϊ��ͨ�ñ��벻ͬ�ı��몤
����������һ��������Ϊ��һ�ְ�������룬���������백����֮��Ķ�Ӧ��ϵ����60����о�����ˡ�1964�� Nirenberg��һ��RNA�ۺ�ø����ϳ��˶�������ᡢ���������ȶ�ۺ����ᣬ����Щ��ۺ�����ֱ����ڵ����ʵ�����ϳɡ����֣������õĶ�ۺ� ����Ϊ���������ʱ��ֻ�ж�۱�������ϳɣ�����ζ��UUUΪ����������룻��������ۺ����������Ӧ��ʵ����֣�CCCΪ��������룬��AAAΪ���� ����룻����������ú��������Ϊ��֪�����Ǻ�������������Ķ�ۺ����ᣬ�Լ�����֪���еĺ����ֻ��������Ϻ�����Ķ�ۺ����������Ӧ��ʵ�飬����� ��������ͳ�ƴ������ֽ����һ�������ӣ����а���������ֹ�룬�����һЩ��������ͨ���ϳ���֪���е����ۺ�������˵���������з�����ͬλ�ر�ǵİ� �����tRNA������ԭ�����Խ���ġ��������������У�AUG����Ϊ��������룬�������Ƿ���(translation,��mRNA�ϵ��Ŵ���Ϣָ���˵� �����϶������ϳɵĹ���)����ʼ�źţ�UAA��UAG��UGA��Ϊ�κΰ�������룬������Ϊ�������ֹ�źţ�ͳ��Ϊ��ֹ��(stop codon)���ֳ���������������(nonsense codon)��
�������������������һ�����ϵ������������롣�����ʵ�����һ�����⣺����ͬһ�ְ������һ�������ӵ�ʹ��Ƶ�� �Ƿ���ͬ?ϸ�µķ���������������ԭ��������Ǹߵ������������ӵ�ʹ��Ƶ�ʲ�����ƽ���ģ���Щ�����ӵ�ʹ���ʺܸߣ���Щ����ʹ�ã���ʹ��Ƶ�� ��Ҫ��ϸ����tRNA����������ء�
(��)ת��RNA(tRNA)
����tRNA(transfer RNA)�ǵ����ʺϳ��еĽӺ������ӡ�tRNA������100���֣�����Я��һ�ְ����ᣬ����ת�˵��˵������ϣ��������ʺϳ�ʹ�á�tRNA��ϸ���ڷ����� ��С��һ����ᣬ��70��120�����ṹ�ɣ�����tRNA������һ���ṹ�ϣ������ڶ��������ṹ�Ͼ���һЩ��ͬ�ص㡣tRNA�к���10%��20%��ϡ�� ���(rare bases)���磺����������mG��mA��˫�������(DHU)���λ����ʵȵȡ����⣬tRNA�ڻ�����һЩϡ�к��գ��磺������ऺ��Ǻ��գ�������� ����(Ψ��pseudouridine)�ȡ��������һ�������DNA�У��ڼ�����ऺ����У�����ͨ����ऻ���1λ��ԭ�ӣ�������ऻ��е�5λ̼ԭ������ �ǵ�1′λ̼ԭ��֮���γ����ռ���
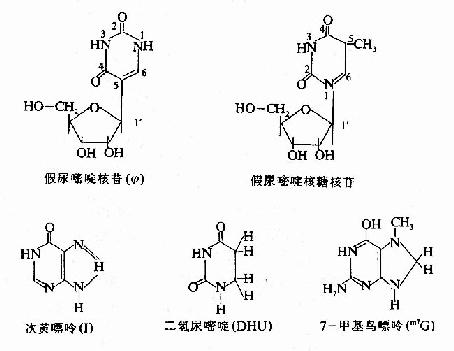
����tRNA�����ڵĺ�����ͨ�������������γɶദ�ֲ�˫�����ṹ��δ��˫����������������ν�Ļ����ᡣ�ַ��� ������tRNA���ɳ���ͼ15-14��ʾ��������ν����Ҷ����(clover leafpattern)�����ṹ���ڴ˽ṹ�У���5′ĩ����ĵ�һ������DHU�����Ժ����������Ϊ�������ڶ�����Ϊ�������ӻ����价�в������������ ����mRNA�е��������������γɼ��������ԣ�������ν�ķ�������(anticodon)���ڵ����ʺϳ������������ӣ�����ȷ�İ���������ϳ�λ��� ���ã���������ΪTΨ�����Ժ����ٺ��պͼ�����Ϊ�������ڷ������ӻ���TΨ��֮�䣬��������һ���ᣬ������������ʮ�����������ɣ�����tRNA3′ĩ�� ������ͬ��CCA-OH�ṹ��tRNA��ת�˵İ�����������ڴ�ĩ���ϡ�(ͼ15��14A)

ͼ15��14��tRNA�Ķ����������ṹ��
A.�����ṹ(aʾ�����뻷��������)��
B.�����ṹ(����ʾ���ܵķdz���������������)
����ͨ��X����������Ƚṹ��������������tRNA�Ĺ�ͬ�����ṹ���ʵ�L��(ͼ15��14B)������3′ĩ�˺� CCA��OH�İ������λ��һ�ˣ��������ӻ�λ����һ�ˣ�DHU����TΨ�����ڶ����ṹ�ϸ���һ�������������ṹ��ȴ��ڽ���tRNA�����ṹ��άϵ�� Ҫ������������֮���γɵĸ������������tRNA���ӵĺ��������кͳ������ϴ��������ṹ�����ƣ���ʾ���ֿռ�ṹ��tRNA�Ĺ��������й�ϵ��
����(��)�˵�����RNA(rRNA)
�����˵�����RNA(ribosomal RNA)��ϸ���ں�������RNA��ԼռRNA������80%���ϣ��ǵ����ʺϳɻ������˵�����(������)(ribosome)����ɳɷ֡������嵰�� (ribosmal protein,rp)����ʮ�֣�����Ƿ���������Ķ����࣬�ֲ��ں˵�������ǻ��ĵ��׳�Ϊrpl����С�ǻ��ij�rps��
����ԭ��������������ĺ˵�����������ڽ�۵Ĵ�С�ǻ���ɡ��Դ˾��˵�������о�����������������֮�� ��rNRA������֮һ�ǵ����ʡ�rRNA��Ϊ5S��16S��23S���֡�S�Ǵ���������ڳ������ij����е�һ������ѧ��λ���ɷ�ӳ�������Ĵ�С��С�ǻ��� 16SrRNA��21��rps���ɣ����ǻ���5S��23s rRNA��31�� rpl���ɡ��������˵�����Сҵ����18S rRNA��30����rps�����ǻ���28S��5.8S��5S����rRNA����50��rpl����������˵�����С�ǻ��е�rRNA�������ƵĶ����ṹ(ͼ 15��15)��
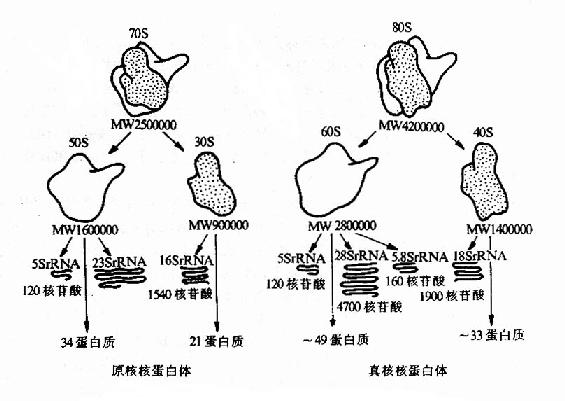
ͼ15��15��ԭ���������������˵�����Ľṹ�ȽϪ�
(������˵�����Ľṹ��ԭ������)
�����������Թ��ڻ�ϸ���ڣ���С�ǻ���������ɺ˵�������������������֡���ʮ�ֶ�������λ������ᣬ������ �뼸��rRNA��������?���ᴿ�˵��ǻ����е��ĺ�rRNA���Թ��ڻ�ϣ����ֲ������ø��ATP�Ϳ����Զ���װ��Ϊ�л��Ե��ǻ�����rRNA֮��ȴ���� ���������Ҳ��˵����������װ��������rRNAΪ�����ġ���Ȼ���ж�������װ��Ҳ��ȱһ���ɵģ�����ͬ���Ŀ�����ø�����û����ЧӦ������֤��ijЩ���� �嵰����ø�Ĺ��ܣ������д������δŪ����������á�
����(��)����RNA����
����С��RNA(snRNA,small nuclear RNA)���������ϸ����ϸ�����ڣ���һ���ΪС�˺˵����帴����(snRNP)����ɳɷ֣���U1��U2��U4��U5��U6snRNA�ȣ���ΪС���Ӻ� �Ǻ��ᣬ��Լ106?89�������ᣬ�书������hnRNA����ת��ΪmRNA�Ĺ����У�����RNA�ļ��ӣ������ڽ�mRNA��ϸ�����˵�ϸ�����Ĺ������� ��ʮ����Ҫ������(��15��10)��
��15��10��snRNA�������빦��
snRNA ���Ӵ�С������������ ���� U1 165 ���5′�����ӵ� U2 185 ����ڷ�֧�� U5 116 �����3′�����ӵ� U4 145106 װ����ӿ��� U6
����С����RNA(scRNA,small cytosol RNA)�ֳ�Ϊ7SL��RNA����Լ300�������ᣬ��Ҫ������ϸ�����У��ǵ����ʶ�λ�ϳ��ڴ�����������������ź�ʶ����(signal recognization particle)����ɳɷ֡�
��
�����������ɺ�����ۺ϶��ɵĸ߷��ӻ���������������Ŵ���Ϣ��Я���ߡ����ݺ�������������ǵ����ͣ��������Ϊ�������Ǻ���(DNA)�ͺ��Ǻ���(RNA)�����ࡣ
����������������������Ǻͺ��������ɣ�����������ʺ���������ࡣDNAһ�㺬A��C��G��T���ּ����RNA��A��C��G��U���ּ����
�������ֺ����ᰴ��һ��������˳��ͨ��3′��5′��������������γɵ����ζ�����ἴDNA��һ���ṹ����ͬ����˳���DNA���ι��ɵ��ض����ܵ�λ������DNA��һ���ṹ�����˻���Ĺ��ܡ�
����һ�㽫ϸ����Ⱦɫ�����DNA�������Ϊ�������� ͬһ���ֵĻ�����DNA�������Ǻ㶨�ģ���ͬ���ּ�������С���ӳ̶Ȳ��켫�����������и��ӵ�Ⱦɫ��ṹ���������DNA�ϴ����ŵ�һ���кʹ��� �ظ����У��������˻����Dz������ģ��ڳ���RNA�г��ֵIJ��ֳ�Ϊ�����ӣ���DNAƴ�ӹ����б�ɾ���IJ��ֳ�Ϊ�ں��ӡ�ԭ������û�к�Ĥ����DNA�� RNA�͵�����һ���γ�һ����Լ��е�������ˡ�ԭ������������Ϲ�����صĻ���������һ��ת¼��ͬһmRNA�����У��γɶ�˳���ӽṹ��ijЩ�� ���л���ֻ����ص���
�����ڽ���������DNA���ܷ���ͻ�䣬��Ӱ����������͵�DNAͻ���Ϊ����ͻ�䣬����ͻ�䳣���ϵ³������Ŵ���ʽ�Ŵ�����һ�������е�������Ƭ�γ��ȶ�̬���ѱ��㷺�����Ŵ�������ϡ���ǰ��ϡ����Ӽ����Լ���ҽѧ�϶��ﷸ��ȷ�ϵȡ�
����˫�����ṹ��DNA�Ķ����ṹ�������Ǻ���������ɵ����������Է�ƽ�еķ�ʽ�����ַ�������ƣ�����˫���� �ĹǼܡ�������������ˮ�Զ�����˫����������棬���������һ���̶ȵ���ˮ�Զ�λ��˫�������ڲ����������ϵļ����A��T��G��C�Ļ���������������� �ӣ������Ŵ���Ϣ�ɿ����ݡ�DNA�뱣�����ƵĻ���������������������˫�����Ŀռ䣬���ڱ����γɴ�С�������ǵ��ص�����ʶ��DNA��Ϣ����Ҫ�� ����
����DNA���ӽṹ����һ�ɲ��䣬�����ڲ�ͬ�����¿���������ͬ����DNA�ṹ�Ķ�̬�ԡ�������״̬��DNA��ҪΪB����������������A�����Z����Z������Ψһ���ڵ�����˫��������
����DNA���ϵ����ּ��Ҳ�Ǿ��ȷֲ������������һЩ��������С��������������������Ժ�������ø�͵��ص���ʶ��λ�㣻����A/T����������������Ŵ�ѧ����������ȱ�ٵģ����ʺ���व�����˳���˫�������ȶ���Ҳ���ش�Ӱ�졣
����DNA���Ȼ��������Լ��������£�˫�����ṹ��ƻ���˫���������룬�����ԡ�����DNA��ijЩ�������ʺ��� ��ѧ������֮�ı䣬����ɫЧӦ��������ȱ��Թ��������������ֵ�ﵽ���ֵ��50%ʱ���¶ȳ�Ϊ����Ľ����¶�(Tm)��Tmֵ�Ĵ�С�������Ӵ�С�� G��C��ռ�ܼ�����İٷֱȳ�����ء����Ե�DNA�������ʵ����������ָܻ�˫�����ṹ�����������á���ͬ��Դ�ı��Ժ���һ���ԣ�����ܷ����ӽ����ӽ� �������������ѧ�����Ļ�����
����˫����DNA��һ��Ť�����ɵij�������ΪDNA�������ṹ����������У�DNA���鵰���γɺ�С��ṹʱ�������Ÿ�����������С���ǹ���Ⱦɫ�ʵĻ����ṹ��λ��ÿ����С�嵥λ����200bp���ҵ�DNA��һ���鵰�װ˾����Լ�һ�����ӵ��鵰��H1��
����RNA����mRNA��hnRNA��rRNA��tRNA��snRNA��scRNA�����Ǿ����Ŵ���Ϣ�ı����� �ء�mRNA���Ŵ���Ϣ��Я���ߣ�����������о����źϳɵ����ʵİ��������У�hnRNA��mRNA��ǰ�壬����ת¼�ġ����������ڳ���mRNA�еĺ��� ��Ƭ��(�ں���)��tRNAʶ�������ӣ�����ȷ�İ�����ת���������ʺϳ�λ�㣻rRNA�ǵ����ʺϳɻ���——�˵��������ɳɷ֣�snRNA��hnRNA ��mRNAת����̵ļ�������ʮ����Ҫ�����á�
��һƪ��DNA������ϳ�
��һƪ��DNA�������ṹ�빦��










