DNA������ϳ�
2012-01-26 11:06:13 ��Դ��37��ҽѧ�� ���ߣ� ���ۣ�0 �����
�������Ǻ���(DNA)��������Ŵ�����Ҫ���ʻ����������л�����Ŵ�����������(code)����ʽ������DNA�����ϣ� ����Ϊ�ض��ĺ���������˳��-���Ŵ���Ϣ����ϸ������ǰͨ��DNA�ĸ���(Replication)�����Ŵ���Ϣ���״����ݸ��Ӵ����ں���ĸ��巢������ �У��Ŵ���Ϣ��DNAת¼(Transcription)��RNA����ָ���������ϳɣ���ִ�и����������ܣ�ʹ������ֳ����״����Ƶ��Ŵ���״�������Ŵ� ��Ϣ�Ĵ��ݷ����Ǵ�DNA��RNA�ٵ������ʣ�����ν������ѧ“���ķ���”��80����Ժ���ijЩ�°�RNA�����з����Ŵ���ϢҲ�ɴ�����RNA�����У� ��RNAͨ����ת¼(reverse transcription)�ķ�ʽ���Ŵ���Ϣ���ݸ�DNA����Ϊ���ķ���������µ����ݡ�Ŀǰ��Ϊ������Ŵ���Ϣ���ݵ����ķ���Ϊ��
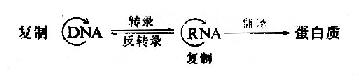
�������µ�������Ҫ�漰DNA����ϳɵ��������棬��һ��DNA���ƣ��ڶ���RNA��ת¼ΪDNA��������ϸ����DNA�ܵ�����ʱ���е������á�
��һ�ڡ�DNA�ĸ���
����DNA��Ϊ�Ŵ����ʵĻ����ص������ϸ������ǰ����ȷ�����Ҹ���(self��replication)��ʹ DNA�����ɱ����ӣ�����ϸ�����ѵ����ʻ�����1953��Watson��Crick���DNA˫�����ṹģ��ָ����DNA���ɶ����������������������� �ɣ�����һ��DNA���ϵĺ���������˳������ͼ16-1˫����DNA�ĸ�����һ�������ġ����˵��DNA�ĸ�������ԭ�����ڵķ���Ϊģ�����ϳ��µ������� ���й����ֹ���DNA���Ʒ�ʽ��ѧ˵�������뱣�����ƣ�ȫ���������Լ���ɢ���Ƶ�(ͼ16-1)��
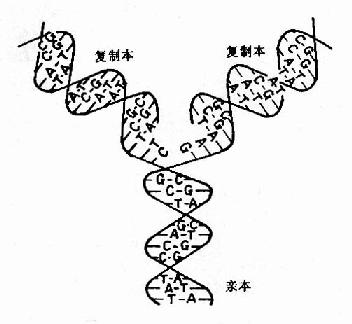
ͼ16��1��˫����DNA�ĸ���
����һ��DNA���Ƶķ�ʽ��һ����̣�
����(һ)DNA�İ뱣������(semiconservative replication)
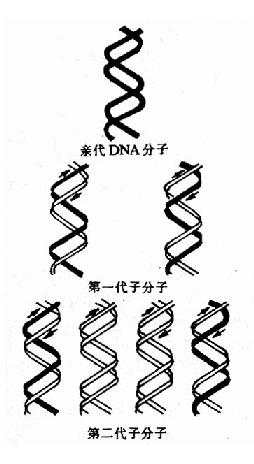
����Watson��Crick�����DNA˫�����ṹģ��ʱ���Ʋ⣬DNA�ڸ���ʱ����������֮�������������� ���ֿ���Ȼ����ÿһ�����ֱ���ģ����Ժϳ�һ���µ�DNA���������ºϳɵ��Ӵ�DNA������һ���������״�DNA����һ�������ºϳɵģ����ָ��Ʒ�ʽΪ�� �������ơ�
����1958��Meselson��Stahl���õ���Ǽ����ڴ˾����״�֤ʵ��DNA�İ뱣�����ƣ����ǽ��� ���˾����ں���15N��ǵ�NH4Cl�������з�ֳ��15����ʹ���еĴ˾�DNA��15N����ǣ����Եõ�15N�DNA��Ȼ��ϸ��ת�Ƶ�����14N ��ǵ�NH4Cl�������н�����������������ͬ����ʱ���ռ�ϸ�����ѽ�ϸ�������Ȼ��(CsCl)�ܶ��ݶ����ķ��۲�DNA������λ�á�����15N�DNA ���ܶȱ���ͨDNA(14N��DNA)���ܶȴ����Ȼ���ܶ��ݶ�����(density gradient centrifugation)ʱ�������ܶȲ�ͬ��DNA�ֲ��ڲ�ͬ��������
����ʵ������������ȫ����15N��ǵ��������еõ���15N�DNA��ʾΪһ�����ܶȴ�λ�����Ĺ��� �ܵס���ת��14N��ǵ��������з�ֳ���һ�����õ���һ�����ܶȴ�������15N�DNA��14N-DNA���ӽ����ӡ��ڶ��������ܶȴ������ܶȴ������� ������������Ƿֱ�Ϊ15N14N��DNA��14N14N-DNA�������Ժ���14N���������������������ӣ����ܶȴ���ǿ�������ܶȴ����������Ľ� ���ӹܵ��ܿڣ�CsCl��Һ�ܶȷֲ��Ӹߵ����γ��ܶ��ݶȣ���ͬ������DNA���Ӿ�ͣ���������൱��CsCl�ܶȴ�����������¿��Կ���DNA���� �γɵ�������Ϊ��֤ʵ��һ���ӽ�����ȷʵ��һ��15N-DNA����14N��DNA���������ӽ����Ӿ����ȱ��ԣ����ڱ���ǰ���DNA�ֱ����CsCl�ܶ� �ݶ����ģ��������ǰ���ӽ�����Ϊһ�����ܶȴ������Ժ����Ϊ���������������ܶȴ�(15N��DNA)�����ܶȴ�(14N��DNA)�����ǵ�ʵ��ֻ���ð� �������Ƶ����۲��ܵõ�Բ���Ľ���(ͼ16��2��16-3)��
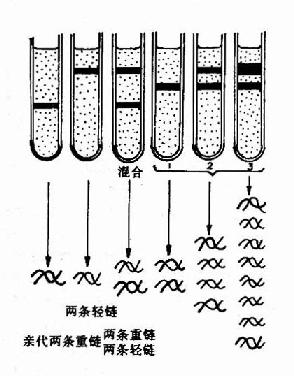
ͼ16-2��DNA�İ뱣�����Ƶ�һ������
����һ���״�����(�ú�ɫ��ʾ)������
һ���ºϳɵ���(�ð�ɫ��ʾ)��ԡ���
�Ժ���������ƹ����У�ԭ���״�����
������Ȼ�������������������������
������һ��ԭ���״�������
ͼ16-3��DNA�İ뱣����
�ƣ�Meslson��Stahlʵ��
�ܶ��ݶ����ĺ��DNAλ
�ã�������Ϊ���գ�����
��Ϊʵ����
������
����(��)DNA���Ƶ�һ����̣�
����DNA˫�����������������෴�ĵ�����ɣ����ƿ�ʼʱ��˫�����γ�һ�����Ʋ�(replicative fork,�Ӵ������һ�������γ�)��һ��������(replicative bubble���Ӵ���������������γɡ�)���������ֱ���ģ�塣���Ժϳ�һ���µ�DNA��������DNAһ������������5′→3′������һ���������� ��3′→5′������������DNA�ۺ�øֻ�ܴ�DNA��5′→3′�ķ���ϳɡ���ô����������ͬ��������������ģ����?����������ձ�ѧ�߸������� �����
����ԭ��������3′→5′�����ĸ��Ϊģ��ʱ�����ƺϳɳ�һ��5′→3′�����ǰ���� (leadingstrand)��ǰ������ǰ�������븴�Ʋ������һ�µģ����ǰ�����ĺϳ����������еģ�����һ��ĸ��DNA��5′→3′�������� Ϊģ��ʱ�����ƺϳ�������5′→3′����Ķ��������������(lagging strand)���������ǰ���������븴�Ʋ�Ĵ����෴�ġ������ֻ������Ƭ�ε���ʽ�ϳɣ���ЩƬ�ξͽ�������Ƭ��(Okazaki fragments)��ԭ���������Ƭ�κ���1000-2000�����ᣬ�������һ��100?00�����ᡣ����ٽ��������Ƭ�����ӳ�һ���������������� ǰ�����ĺϳ����������еģ���������ĺϳ��Dz��������еģ����Դ������Ͽ�DNA�ĸ����ǰ벻��������(ͼ16-4)��
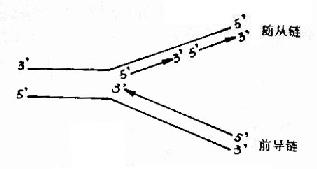
ͼ16��4��DNA�İ벻��������
����DNA���Ƶ�ȫ�����̿�����Ϊ�طֳ������Σ���һ����ΪDNA���Ƶ���ʼ�Σ�����ΰ�����ʼ�㣬���� �����Լ���������γɣ��ڶ���ΪDNA�����ӳ�������ǰ��������������γɺ��г�RNA��������ȱ�����Ӹ���Ƭ�Ρ�������ΪDNA���Ƶ���ֹ�� �Ρ���DNA���Ƶ�������������Ҫ30����ø�������ʷ��Ӳμӣ����ǽ���DNA���Ƶĸ����������ؽ������ǵ����á�
��������DNA���Ƶ���ʼ�Σ�
����(һ)DNA���Ƶ���ʼ��
�����ܶ�ʵ�鶼֤���������Ǵ�DNA�����ϵ��ض���λ��ʼ�ģ���һ��λ����������ʼ��(originof replication)����ori��o��ʾ��ϸ���е�DNA����һ����ʼ�ͻ�����������ȥ��ֱ�����ϸ����ȫ��������DNA �ĸ��ơ�DNA���ƴ���ʼ�㿪ʼֱ���յ�Ϊֹ��ÿ��������DNA��λ��Ϊ�����ӻ��Ƶ�Ԫ(replicon)����ԭ��ϸ���У�ÿ��DNA����ֻ��һ���� ����ʼ�㣬���ֻ��һ�������ӣ�������������У�DNA�ĸ����Ǵ�������ʼ��ͬʱ��ʼ�ģ�����ÿ��DNA������������������ӡ�
����DNA������ʼ���нṹ�ϵ������ԣ����磺�˾�Ⱦɫ��DNA������ʼ��Oric��422����������ɣ��� һϵ�жԳ����еķ����ظ����У������Ľṹ(palindrome)��������9���������13����������ɵı������У���Щ��λ�Ǵ˾���DnaA���� ʶ���λ�ã��˾�Ⱦɫ��DNA�ǻ�״˫��DNA�����ĸ����ǵ��͵�“θ”����(������״��ϣ����ĸθ)����һ����㿪ʼ��ͬʱ������������и��ƣ� ���������Ʒ�������ʱ�����ƾ�ֹͣ������Щ�����DNA������ʼ����һ�θ���A·T�����Ρ���Щ����Ľṹ������DNA������ʼ�����в����ø�����൰�� �ʷ��ӵ�ʶ��ͽ�϶��DZ���ġ�
����(��)DNA���Ƶķ���
����(1)���㿪ʼ˫���ƣ�
��������ԭ��������������DNA������
��Ҫ����ʽ����һ���ض�λ����������������෴�ķ�������������������γ�һ�������ݣ��õ����������Թ۲쵽�����ݵĴ���(ͼ16��5)��

��ͼ16��5��SV40DNA�������������ĵ羵ͼ��
����(2)���㿪ʼ�����ƣ�
��������colE1�Ǹ����͵����ӣ����ƴ�һ����ʼ�㿪ʼ����ͬһ�������������������γ�һ�����Ʋ�(replication fork)��
����(3)���㿪ʼ�����ƣ�
�����ٲ���DNA�ĸ����Ǵ�������㿪ʼ�ģ��γ��������Ʋ棬����һ����һ�����Ƴ�һ������(ͼ16��6)��
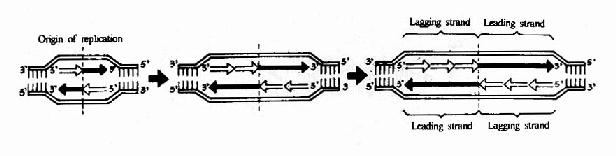
ͼ16-6��DNA�İ벻�������ƺ����ݵ��γ�
������֮DNA���Ƶ���㼰����ԭ��ϸ�������ϸ����ͬ������ͬ����ԭ��������������IJ�ͬ����Ҳ���൱��IJ���(ͼ16��7)��

ͼ16-7��DNA��������������ֻ���
����(��)DNA������ʼ��������γɼ��������ø�͵����ʣ�
����1.����ø(helicase)
����DNA��ʼ����ʱ��������ʼ�㴦�˫������Ӧ����һ�ֽ���ø(helicase)�Ĵ��½��еġ�����ø�� ҪATP�ֽ���������˾���DnaB�����н���ø���ԣ����������ģ��DNA��ϣ���5′→3′�����ƶ�������һ�ֽ���Rep����ǰ������ģ�� DNA�����3′→5′�����ƶ�������ø�����þ��Ǵ�DNA˫��֮��������
����2.������ϵ��ף�(single strand binding proteins,SSBP)
���������ĵ���DNA��ϣ�ʹ���ȶ������ٶ����������ұ����������ø�Ե���DNA��ˮ�⣬��֤�˵���DNA��Ϊģ��ʱ����չ״̬��SSBP�����ظ�����(ͼ16��8)��
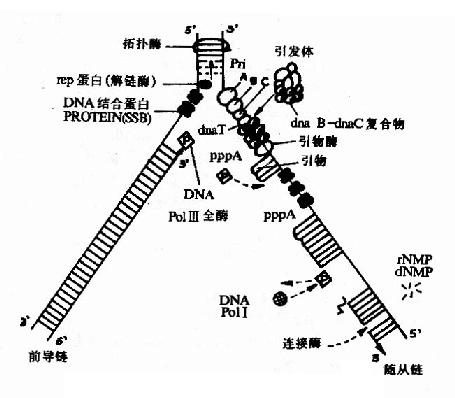
ͼ16-8���˾�DNA���Ʋ��и��ƹ��̼�ͼ
����3.��������γɣ�
����DNA������ʼ�Ĺؽ�������ǰ����DNA�ĺϳɣ�һ��ǰ����DNA�ľۺ����ÿ�ʼ�������DNA�ĺϳ�Ҳ���� ��ʼ������ǰ�����ĺϳ����������еģ�����������ʼ��Լ���������ĺϳ��Dz��������еģ����������αȽϸ��ӡ��˾�������ǰ����Dna B. Dna C�͵�����ϵ�����ɡ�
����(1)����ø(primase)
��������һ�������RNA�ۺ�ø���ɴ���Ƭ��RNA�ĺϳɡ����ֶ�RNAƬ��һ��ʮ��������ʮ��������ȣ�������DNA������ʼ����Ϊ���RNA�����3′�OHĩ���ṩ����DNA�ۺ�ø���γ�DNA���ӵ�һ�������������λ�á�
����(2)������(primosome)
�����߶Ƚ�����ģ��DNA����ֵ����������γɵ�����ǰ��ٽ�����ø�����������ͬ�γ������壬��������Ҫ�� DNA������Ͽ�ʼ����������������ø��ϲ����룬�Ӷ��ڲ�ͬ��λ��������ø���ϳ�RNA���������RNA��3′�OHĩ�˽���ȥ�ϳ�DNAƬ�Σ���� ��������������ϳɵĿ�ʼ��
��������DNA���Ƶ��ӳ����Լ������ø�͵����ʷ��ӣ�
����DNA�ĸ���ʵ���Ͼ�����DNAΪģ����DNA�ۺ�ø�����£������������������������(dATP,dGTP,dCTP,dTTP,��дΪdNTP)�ۺϳ�DNA�Ĺ��̡�
��������һ���dz����ӵ�ø�ٷ�Ӧ����Ҫ������ø�͵����ʲ��룬�ֱַ�����������DNA���������á�
����(һ)DNA�ľۺϷ�Ӧ��DNA�ۺ�ø

ͼ16��9��DNA�ۺ�ø������
����1957�꣬Arthur kornberg�״��ڴ˾��з���DNA�ۺ�ø��(DNa polymerase ��,��дDNA pol��)��������̷�����DNA�ۺ�ø���DNA�ۺ�ø��(DNa polymerase ��дDNA pol��,DNA pol��)ʵ��֤���˾���DNA���Ƶ���Ҫ���̿�DNa pol�������ã���DNA pol���DNA pol����DNA�����У�������������á�����16��1��
��������ø�Ĺ�ͬ�����ǣ�����ҪDNAģ�壬�������ø�ֳ�Ϊ����DNA��DNA�ۺ�ø(DNa dependent DNA polymerase, DDDP)������ҪRNA��DNA��Ϊ����(primer)����DNA�ۺ�ø���ܴ�ͷ��DNA����ʼ���۴�dNTP�ӵ������3′�OHĩ�ˣ���� DNA�ϳɵķ�����5′→3′��ͼ16-9��������DNA�ۺ�ø�����ڶ��ø��������DNA���ƺ������̵IJ�ͬ�η������á�����DNA�ۺ�ø������ ������������Ҵ���������DNA�ۺ�ø�Ļ����ص㣬�����������ؽ���DNa pol������ò�ָ���������DNA pol�������ԣ�
����1.DNA�ۺ�ø��
����DNA pol������һ����������ɣ�������Ϊ109KD��ø�����к���һ��Zn++���Ǿۺϻ��Ա���ġ�
�����˾�ÿ��ϸ����Լ��400��ø���ӣ�ÿ��ø����ÿ������37�����ܴ�667����������뵽DNA���У��ÿݲݸ˾�����ø�ɽ���øˮ�������Ƭ�Σ���Ƭ�η�����Ϊ76KD��ͨ����ΪklenowƬ�Σ�СƬ��Ϊ34KD����СƬ�ξ��в�ͬ��ø���ԡ�
����(1)DNA�ۺ�ø��5′→3′�ۺϻ��ԣ�
��������DNA�ۺ�ø����Ҫ�Ļ��ԣ���ģ��DNA�ϵĺ�����˳��������dNTP����ӵ�����RNA3′�OHĩ �ˣ����ٽ�3′�OH��dNTP��5′�PO4�γ������������ø��רһ�Ա���Ϊ�½����dNTP������ģ��DNA������ʱ���д����ã�5′→3′�ۺϻ� �Դ�����klenowƬ����(ͼ16��9��ͼ16-10)��
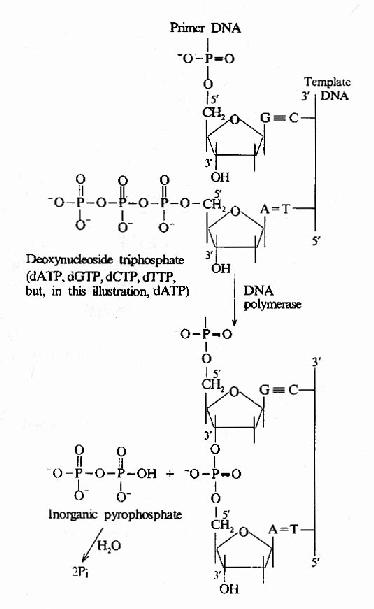
ͼ16-10��DNA�ۺ�ø����DNA���ӳ�
����(2)DNA�ۺ�ø��3′→5′���к���ø���ԣ�
��������ø���Ե���Ҫ�����Ǵ�3′→5′����ʶ���г�DNA������ĩ����ģ��DNA����Զ�����ĺ����ᣬ���ֹ��ܳ�ΪУ�Թ��ܣ����DZ�֤��ۺ����õ���ȷ�Բ���ȱ�ٵģ���˶���DNA�����м��ߵı�������������Ҫ�ġ�
����(3)DNA�ۺ�ø��5′→3′���к���ø���ԣ�
��������ø�����Ǵ�DNA����5′����3′ĩ��ˮ������Եĺ����ᣬ�������ж������������ÿ�����г�10�������ᡣ��ˣ�����ø������DNA���˵����п�������Ҫ���ã�����ɵ�DNAƬ��ȥ��5′�˵�RNA����Ҳ�DZ���ġ�
����DNA pol���5′→3′�ۺϻ��Ժ�5′→3′����ø����Эͬ���ã�����ʹDNAһ�����ϵ��пڴ�5′→3′�����ƶ������ַ�Ӧ����ȱ��ƽ��(nick translation)�����ô˷�Ӧ���������DNAƬ�ν��з�������(α-32PdNTP)�ı���Ƴ�̽��(probe)�����к���ķ����ӽ�ʵ�飬 ���ִ���������ѧ��һ����Ҫ����(ͼ16��11)��
��
��ͼ16-11��ȱ��ƽ�Ʊ��DNA̽��
��������ʵ��֤ʵDNA pol����DNA���ƹ����е���Ҫø������������Ҫ��DNA���˺�����йء�
����2.DNA�ۺ�ø��(DNA pol��)
������ø������Ϊ120KD��ÿ��ϸ��Լ��100��ø���ӣ�������ֻ��DNa pol���5%��������5′→3′�ۺϻ��Ժ�3′→5′���л��ԣ���û��5′→3′���л��ԣ��������ÿ�����DNA�������йء�
����3.DNA�ۺ�ø��(DNA pol��)
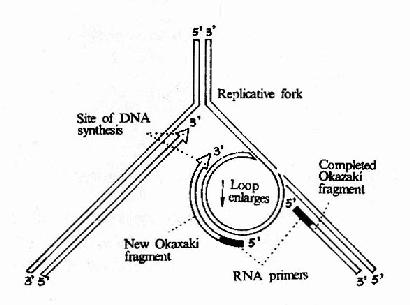
ͼ16��12��DNA�ۺ�ø����ȵ�������ӵĺϳ�
����������DNA���ƹ���������Ҫ���õľۺ�ø��������һ�����ǻ���ɵĵ����ʷ��ӣ����������600kDa���� ø�����γ�һ�����ԳƵĶ����壬ÿ���˾�ϸ����ֻ��10?0��ø���ӣ�����dNTP����DNA��������ȴ�����ģ�ԼΪ9000������/ÿ����/ÿ ��ø���ӡ���Ҳ֤��DNa pol����DNA���ƹ�������Ҫ�������õ�ø
���ڴ˾�Ⱦɫ��DNA�� �и���ʱ��DNA�ۺ�ø��ȫø�����ǵ��������õģ������������壬����ø�ȹ���һ��������(replisome)�����ڸ�����Ĵ��ڣ��ȵ�������������� ͬʱ���ơ�DNa pol�����ɶ��ǻ���ɵIJ��Գƶ����壬������ͬʱ�����ȵ�����������ĸ��ƣ���φ×174�ĸ����й۲쵽���������ǰ�����DNA����(loop)�Ĵ� �ڡ�ͼ16?2���Կ����������������ģ��DNA��DNA�ۺ�ø��ȫø����ת��180°���γ�һ�����ۣ���˸���Ƭ�εĺϳɷ����ܹ����ȵ����ĺϳɷ����� ���������ƶ�����һ�¡�
��������DNA pol����ǰ�ƶ����ȵ����ĺϳ����ӳ���ͬʱ������Ƭ��Ҳ�ڲ����ӳ�����һ����Ҳ�ڲ�����������Ƭ�κϳɵ�ǰһ��Ƭ�ε�5′��ʱ����һ�����۾��ͷ� ���������ڸ��Ʋ���ǰ�ƶ��ֿɽ���һ�����������ģ���û���������������ϳ��µ����Ȼ�����γ�һ��С�����ۣ������µĸ���Ƭ�εĺϳɡ��ɴ�ģ�Ͳ��ѿ� ����������ĺϳ���Ҫ�����Ե������������ϳɽ���������ǰ�������һ������Ƭ�εij��ȡ�����Ƭ����ɺ���5′�˵�RNA������DNa pol���5′→3′����ø�����г����ɴ���ɵĿ�϶����DNA pol���5′→3′�ۺϻ��Դ�dNTP�õ��������DNA�ĸ�������DNa pol���DNA pol�����������ɵġ�
���������б�˵�����ִ˾�DNA�ۺ�ø�����Ԫ�
��16��1���˾�DNA�ۺ�ø������
�� DNA�ۺ�ø�� DNA�ۺ�ø�� DNA�ۺ�ø�� ������ 109KD 120KD ��600KD ÿ��ϸ���еķ����� 400 17-100 10-20 5′→3′�ۺϻ��� + + + 37��ת���ʺ���������ø����·���� 600 30 30,000 5′→3′���л��� + - - 5′→3′���л��� + + + �п�ƽ�ƻ��� + - - ��dNTP���� �� �� �� ���� �� ���� ���� ȥ������ ���ȱ�����������DNA�ۺ�ø
�����������DNA�ۺ�ø��α��β��γ��δ��ε�����ǵĻ������������ڴ˾�DNA�ۺ�ø������Ҫ�����Ǵ�dNTP��5′→3′�ۺϻ��ԣ�������������16��2����
��16-2���������DNA�ۺ�ø
�� α β γ δ ε �ǻ��� 4 4 4 2 5 ��������KD) ��250 36-38 160-300 170 256 ϸ���ڶ�λ �� �� ������ �� �� 5′→3′�ۺϻ��� �� �� �� �� �� 3′→5′���л��� �� �� �� �� �� ���� ���ơ����� �� ���� ���� �����������ϸ����DNA����������Ҫ���õ���DNA polα����Ҫ����Ⱦɫ��DNA�ĸ��ơ�DNa polβ��ģ���������Ǿ���ȱ�ڵ�DNA���ӣ�����Ϊ����DNA���йء�DNa polγ��������DNA�ĸ����������á�DNA polδ������5′→3′�ۺϻ��ԣ����һ�����3′→5′����ø���ԣ�����Ϊ�������DNA��������DNa polα��DNA polδЭͬ�����½��еģ�ǰ�����ĺϳɿ�DNA polδ�������һ���Ҫһ��ϸ�����ڵ������ӗ��ֳϸ���˿��?proliferating cell nucleus antigen, PCNA)���롣��������ĺϳɿ�DNA polα������ø���������ɡ�
����(��)�볬�����ɳ��йص�ø��
����DNA���ƴ���ʼ�㿪ʼ��һ��������ʱ���ֲ���DNA˫���������Ҫ������ø�����ã���ĵ�������Ҫ������ϵ��������ϣ��ڸ��Ʋ���ǰ�ƶ�ʱ�����ǰ��DNA���������������������������������칹ø�����������ֱ�������ǵ����á�
���������칹ø(topoisomerase)��һ��ı�DNA�������ʵ�ø��������ɴ�DNA�ĸ��������칹 ����Ӧ�����������������ǿ��ܲ�����DNA�ĸ�����ת¼����DNA����ʱ�����Ʋ��н���ǰ��DNA���Ӳ��ֲ�������������������ø���ɳ۳������������ڸ� �Ʋ��ǰ����DNA�ĺϳɡ�DNA������ɺ�����ø�ֿɽ�DNA�������볬������ʹDNA���ơ��۵���ѹ�����γ�Ⱦɫ�ʡ�DNA�����칹ø�Т��ͺ͢� �ͣ����ǹ㷺������ԭ�����P��������С���16��3
��16��3���˾�����������е������칹ø��
���� ���� �Գ����������� ���������칹ø �˾� �п�һ��DNA�� �ɳ۸������� ������� �п�һ��DNA�� �ɳ������������� ���������칹ø �˾� �п�����DNA�� �������������� ����ATP ���븺�������� �� ����� ������� �п�����DNA�� �ɳ��������� ����ATP ���������븺���������������칹ø��(Topo��)����Ҫ�����ǽ���״˫��DNA��һ�����п�һ���ڣ��пڴ�����ĩ���������ᰴ���� �۳������ķ���ת����Ȼ���ٽ��пڷ����������ʹDNA���Ʋ��ƶ�ʱ�������ǰ��DNA���������õ����⣬����DNA���Ʋ������ǰ�������칹ø��� ���������⣬�Ի�״����DNA���д��������ã��Ի�״˫��DNA�Ļ���������Լ�ʹ��״����DNA�γɻ�״˫��DNA��������(ͼ16��13)��
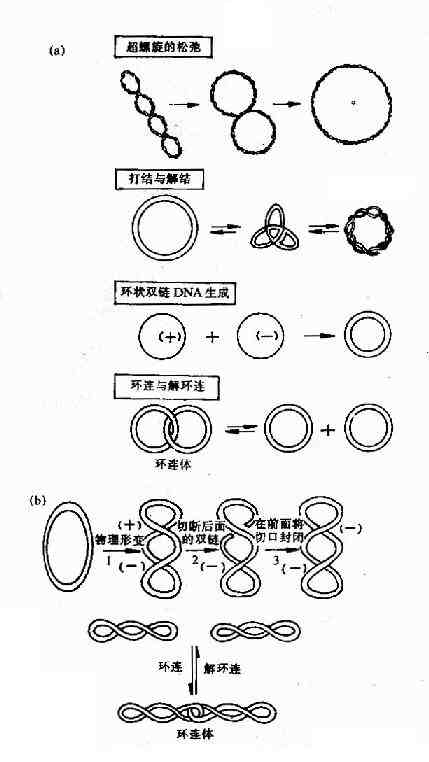
ͼ16����3������ø��������ص�
��(a)�˾�����ø�����4�������칹�����á�(b)����ø�������
���������칹ø��(Topo��)���ڴ˾��з��ֵģ�������Ϊ��תø(gyrase)�����������ص����п���״ ˫��DNA���������������еIJ��־��пڴ�������ת��Ȼ�����пڣ�Topo��ʹDNA���Ӵӳ�����״̬ת��Ϊ�ɳ�״̬���˷�Ӧ����ҪATP���롣 DNA������ɺ�Topo����ATP�����£�DNA���Ӵ��ɳ�״̬ת��Ϊ�������������⣬Topo����������칹����Ӧ���л����������Լ����� ��ᡣ
�����ġ�DNA���Ƶ���ֹ��
����DNA�ڸ��ƹ����У��ϳɳ���ǰ����Ϊһ�������ij���������������ɺϳɳ��������ڵ�Ƭ�Σ�������ø�Ĵ��£����ӳ�Ϊһ������������������������ø���½��еġ�
��������ø(ligase)�������Ǵ����ڵ�DNAƬ����3′��5′����������������ӡ����ӷ�Ӧ�е���������ATP(��NAD��)������ø����ATP���ã��Թ��ۼ���������E�AMP�м������м�������һ��DNAƬ�ε�5′�������������γ�E-AMP-5′��DNA��Ȼ��������һ��DNAƬ�ε�3′-OHHĩ�����ã�E��AMP���£�����DNAƬ����3′��5′��������������ӡ�������ĸ���DNAƬ�ξ����������ӳ�һ��DNA����(ͼ16-14)����
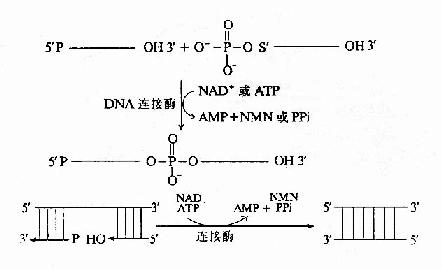
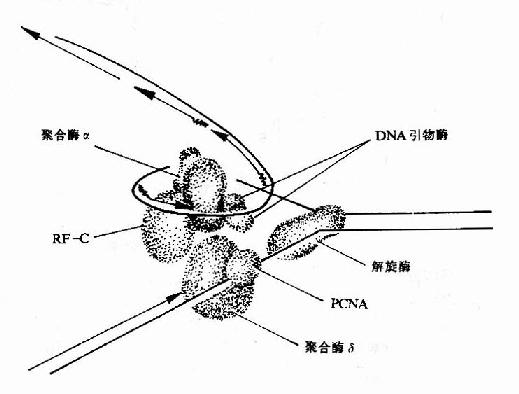
ͼ16-14������ø�Ĵ���Ӧ
ͼ16��15���������DNA���Ʋ�ṹʾ��ͼ
���������о�֤���˾�Ⱦɫ��DNA���и�����ֹλ�㣬�˴����Խ��һ������ĵ����ʷ��ӽ���Tus����������ʿ�����ͨ����ֹ����ø(Helicase)�Ľ������Զ���ֹ���Ƶġ���ϸ�Ļ��ƻ�����ȫ�����
����DNA������ɺ�����ø��DNA�������볬�����ṹ��
�����塢�������D
NA���Ƶ��ص㣺
����DNA���Ƶ��о��������ԭ�������н��еģ���Щԭ�������DNA�����Ѿ���ú��������������ԭ�����︴�ӵö࣬��DNA���ƵĻ������̻������Ƶġ�������������Ҫ����һЩ��Ҫ������
ͼ16��16������ø������TG���ĺϳ�
����1.��ԭ�����ﲻͬ���������DNA������������ʼ�㣬������ĸS.cerevisiae��17��Ⱦɫ��Լ��400����ʼ�㣬��ˣ���Ȼ�������DNA���Ƶ��ٶ�(60������/ÿ����)��ԭ������DNA���Ƶ��ٶ�(E.coli 1700������/ÿ����)���ö࣬��������ȫ��������DNAҲֻҪ�����ӵ�ʱ�䡣
����2.SV40����DNA��Ҫ��������ϸ���е�DNA������ϵ����DNA�ĸ��ƣ������˽��������DNA���Ƶ� ����ģ�͡����������DNA���Ʋ洦����Ҫ���ֲ�ͬ��ø��DNA�ۺ�øα(polα)��DNA�ۺ�øδ(polδ)��polα������ø���ܽ�ϣ���DNA ģ�����Ⱥϳ�RNA�������polα�ӳ�DNA�������ֻ��Ի�Ҫ��������C���롣ͬʱ���������ģ���ϵ�PCNA(��ֳϸ���˿�ԭ Proliferating cell nuclear antigen)��ʱ�ͷ���polα��Ȼ����polδ��ϵ�������3′ĩ�ˣ�����PCNA��ϣ������ϳ�ǰ��������������ĺϳɿ�polα����������ø�� �ϲ��ڸ�������C�����£��ϳɸ���Ƭ��(ͼ16��15)��
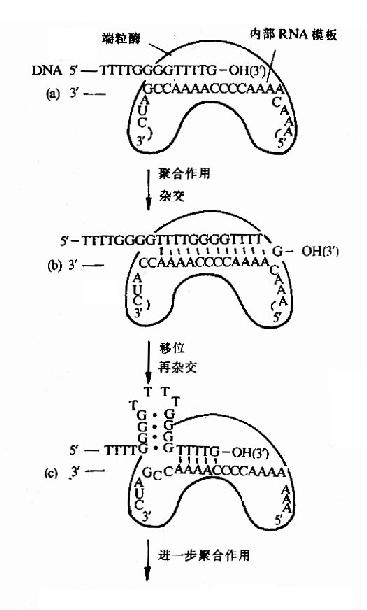
����3.�����������Ⱦɫ��������DNA���������˽�������(telomeres)�����������ظ��ĹѺ��������й��ɵġ�������ĸ�� �����ظ�������5′G(1?)T(3)3′��ǰ�潲����������DNA�ۺ�ø��ֻ�ܴ�DNA��5′→3′�ķ���ϳɣ���˵����Ʋ浽������Ⱦɫ��ĩ��ʱ�� ǰ�������������ϳɵ�ͷ�����������������һ�ֲ���������ʽ�ϳɸ���Ƭ�Σ����Բ����������Ⱦɫ��ĩ�˵ĸ��ƣ����������ⲻ��������������ϸ������ʱ DNA���ƽ�����5′ĩ��������ʹDNA���̣���ʮ������о�����������������ڶ�����һ������ķ�ת¼ø��������ø(telomerase)�������ɵ����ʺ�RNA��������ɵģ�����������RNAΪģ�壬�������ģ��DNA��3′�OHĩ���ӳ�DNA�����������ӳ���DNAΪģ�壬�����ϳ������(ͼ16��16)��
�����ɴ˿ɼ�����ø�ڱ�֤Ⱦɫ�帴�Ƶ�������������Ҫ���塣
��һƪ����ת¼����(reverse transcription)
��һƪ��RNA�Ľṹ�빦��










