Ĥ����鵼���ź�ת��
2011-12-03 17:07:55 ��Դ��37��ҽѧ�� ���ߣ� ���ۣ�0 �����
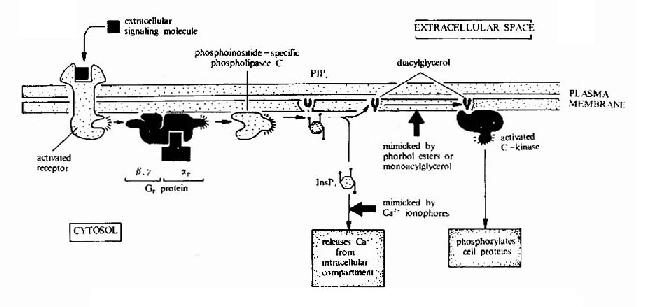
��֬���ԵĻ�ѧ�źŲ�ͬ����ˮ���źŷ���(���е����༤�ء����ʺ���ϸ�����ӵ�)�����ܽ���ϸ�������ǵ�����λ��ϸ �����档��Щ�������źŷ��ӽ�Ϻ����յ�ϸ���ڷ���һϵ�����ﻯѧ�仯���Ӷ�ʹϸ���Ĺ������������ֻ���ϸ���ڻ�ѧ���ʵķֲ��ȷ����ı䣬����Ӧ�� ���ı仯�ͻ���������Ҫ����һ���̿��Գ�֮Ϊ��Ĥ�ź�ת��������һ�ź�ת�������У��źŷ��Ӳ�����ϸ������Ȼ��Щ�źŷ����������Ϻ���Է����ڻ� (internalization)�����ⲻ����Ҫ�����÷�ʽ������λ��Ĥ������������鵼���źŴ�����Ҫ����Ϊ�����ֲ����źŴ��ݵ��źŷ��ӵĹ���Ũ �Ȼ�ֲ������仯�������źŷ���֮�䷢���ʶ�������á�
����һ��Ĥ����ķ���
��������Խ��Խ���Ĥ�������屻��������ṹ��ת���źŵķ�ʽ���Բ�����Ŀǰ����������Ľṹ�������÷�ʽ�ɽ����Ϊ�����ࡣ���������������������ࡢ�����һ��ṹ���ܼ�ϸ����֮������Ӧ�ķ�ʽ��������ͬ������21��2��
Table 21��2��Classification of Membrane Receptors:Characteristics of Three Groups of Receptors
Characteristics Ion Channel
Receptors G-Protein-Linked
receptors Recetpors with a Single
Transmembrane Domain Endogenous ligands Neurotransmitter Neurotransmitter Growth factor hormone Hormone Cytokine Auloacoid Chemotactic factor Exogenous stimulant Structure Oligomer with a pore Probably monomer Monomer of oligoner with (±��catalytic domain Number of transmem Four per subunit Seven One per subunit bane segments Function Ion channel Activation of G proteins Tyrosine kinase Giuanylate cyclase(?) Cellular responses Depolarization or Depolarization or Regulation of function �� hyperpolarization hyperpolarization and expression of Regulation of function proteins and expression of Proliferation or proteins differentiation
��������Ĥ�����ź�ת���ķ��ӻ���
����(һ)����ͨ�������弰���ź�ת��
��������ͨ����������һ������Ϊ����ͨ�������塣��������ͨ�����ܵ�λ���Ƶ�����ͨ�����ܻ�ѧ���ε��ص�����ͨ����ͬ�����ǵĿ��Ż�ر�ֱ��������Ŀ��ƣ���������ҪΪ���ʡ�
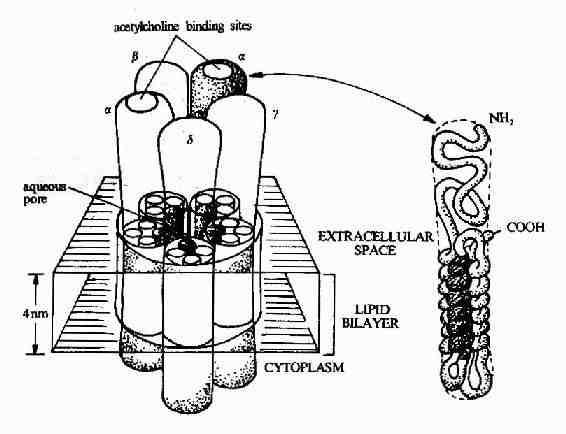
ͼ21��8��������������Ľṹģʽͼ
����ͼ21��8��ʾ����Ϊ����ͨ������ĵ��ʹ�����������������Ľṹģʽ������������������5��ͬԴ�Ժܸߵ��� �����ɣ�����2��α�ǻ���1��β�ǻ���1��γ�ǻ��ĺ�1��δ�ǻ���ÿһ���ǻ�����һ���Ĵο�Ĥ���ף�������Լ60kd��Լ��500��������л����ɡ��Ʋ� ��Ĥ����Ϊ����α�����ṹ������һ��α�������϶�ļ������ᣬ�������������ˮ���Ĵ��ڣ�ʹ����ǻ���ͬ��Ĥ���γ�һ����ˮ�Ե�ͨ������������Ľ�ϲ� λλ��α�ǻ��ϡ�
������������������������ֹ������(ͼ21-9)����������������Ľ�Ͽ���ʹ֮����ͨ�����Ź��� ��ʹ����������Ľ�ϣ������崦��ͨ�����Ź���״̬��ʱ����ʮ�ֶ��ݣ��ڼ�ʮ�������ֻص��ر�״̬��Ȼ������������֮���룬������ָ�����ʼ״̬������ ���½������������
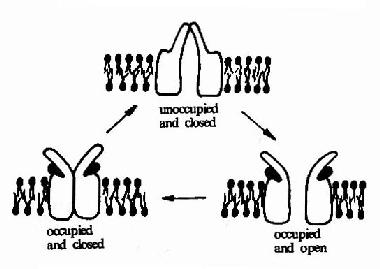
ͼ21��9������������������ֹ���ʾ��ͼ
��������ͨ�������ź�ת�������������ǵ�����ϸ��Ĥ��λ�ĸı䣬������Ϊ������ͨ��������ͨ������ѧ�ź�ת���Ϊ���źŶ�Ӱ��ϸ���Ĺ��ܵġ�
��������ͨ�������������������ͨ��������������Ȱ��������ɫ�������壬Ҳ������������ͨ������ʰ����γ��������������塣
����(��)G����ż�������弰���ź�ת��
����G����ż������������������ʡ����༤�غ��������ӵ����壬��ζ�����Ӿ�������н�����Դ�������ص����� ����G����ż�������塣���������ڽṹ�Ͼ�Ϊ���嵰�ף�����ĩ��λ��ϸ������棬�Ȼ�ĩ���ڰ�Ĥ�ڲࡣ����������Ҫ������Ĥ�ߴ�(ͼ21-10)������� ���˽����������Ϊ�ߴο�Ĥ���塣��������������Ĥ����Ĥ����Ĥ�ڲ��γ��˼�����״�ṹ�����Ƿֱ���������(��ѧ�������ź�)�Ľ�Ϻ�ϸ���ڵ��źŴ� �ݡ���������ֿ�����һ��GTP��ϵ���(���G����)����ã�����G�����Ǹ��źŴ���;���еĵ�һ���źŴ��ݷ��ӣ���Ҳ���������屻��ΪG����ż���� �����ԭ��
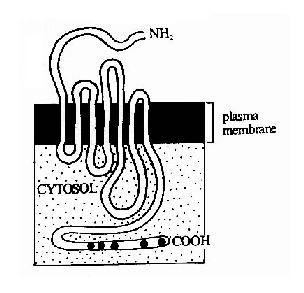
ͼ21��10���ߴο�Ĥ���壭G����ż��
���������Ĥ�ṹʾ��ͼ
����G����ż��������źŴ��ݹ��̰���(1)�����������ϣ�(2)����G���ף�(3)G�����������ϸ�� �е�ЧӦ���ӣ�(4)ЧӦ���Ӹı�ϸ������ʹ�ĺ�����ֲ���(5)ϸ������ʹ��������Ӧ�İз��ӣ��Ӷ��ı�ϸ���Ĵ�л���̼��������ȹ��ܡ����ڽ���һ�� ����һ���̵���Ҫ���ڡ�
����1.G����ѭ����(G Protein Cycle)
����G����ż����������ź�ת��;���еĵ�һ���źŴ��ݷ�����G���ף������̳�ΪG����ѭ����
����G������α��βγ�ǻ����������ʽ������ϸ����Ĥ�ڲࡣα�ǻ��ѷ�����20���֣�������Ϊ36��52kd��α �ǻ����ж���λ�㣬���а������������ϲ��������ڵIJ�λ����βγ�ǻ����ϵIJ�λ��GDP��GTP��ϲ�λ�Լ�������ЧӦ��������õIJ�λ�� �ȡ�α�ǻ�������GTPø���ԡ�α�ǻ����GDPʱ������״̬������GTP���ʱ��Ϊ�л���״̬��GTP��ˮ����ʹ�䷵������״̬��
����G�����е�β��γ�ǻ��������֣�������α�ǻ�����ࡣ��ϸ���ڣ�β��γ�ǻ��γɽ��ܽ�ϵĶ����壬ֻ���ڵ��� ���������·��ɽ��룬��˿�����Ϊ�����ǹ����ϵĵ��塣βγ�ǻ�����Ҫ��������α�ǻ��γɸ����岢��λ����Ĥ�ڲࡣ���������о�������βγ�ǻ���������� ������ЧӦ���ӡ�
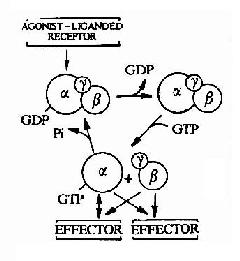
ͼ21��11��G����ѭ��ʾ��ͼ
����G����ѭ���ľ�����̿ɼ�ͼ21-11����������ѧ�źŴ̼�����ʱ������G����ʹ֮��������ı䡣α�� ����GDP�������½�����ϵ�GDPΪGTP��ȡ����α�ǻ������GTP����βγ�ǻ��������룬��Ϊ�״̬��α�ǻ�����˵�α�ǻ���ʱ���������� ���εĸ���ЧӦ���ӡ����ֻ״̬��һֱ������GTP��α�ǻ��������е�GTPøˮ��ΪGDP��һ������GTP��ˮ�⣬α�ǻ����ٴ���βγ�ǻ��γɸ��� �壬�ص���ֹ״̬�����½����µĻ�ѧ�źš�
��������G�������ͬ�����G�����������ڲ�ͬ��ЧӦ���ӣ����ͬһЧӦ���ӽ��в�ͬ�ĵ��ڡ�
����2.ЧӦ���Ӽ�ϸ������ʹ
����G����
�֮�������������ỷ ��ø����֬øC��ЧӦ����(Effector)�ϡ��е�α�ǻ�(Gs)���Լ��������ỷ��ø���е�α�ǻ�(αi)�������������ỷ��ø�������ỷ��ø�� ATP���ɻ�״AMP(cAMP)�ķ�Ӧ�����ϸ���ڵ�cAMPˮƽ�������������Ϻ���G����α�ǵ�λ�����ö����ͣ��Ӷ���ϸ�����ź�ת��Ϊ ϸ�����źš�����ϸ�����źſ������������η��ӡ�����ϸ�����źŵĴ��ݷ�ʽ��G����ż�������崫���źŵ���Ҫ��ʽ����Щϸ�����źŷ��ӱ���Ϊϸ������ʹ�� ϸ������ʹ�౻��Ϊ�ڶ���ʹ����֪��ϸ������ʹ����cAMP��cGMP�����Ͷ���(DAG)��IP3����Ca2���ȵ�(ͼ21��12��13��14)��G�� ��α�ǻ����ࡢ�����õ�ЧӦ���Ӽ������ڵ�ϸ������ʹ�ɲμ���21-3��
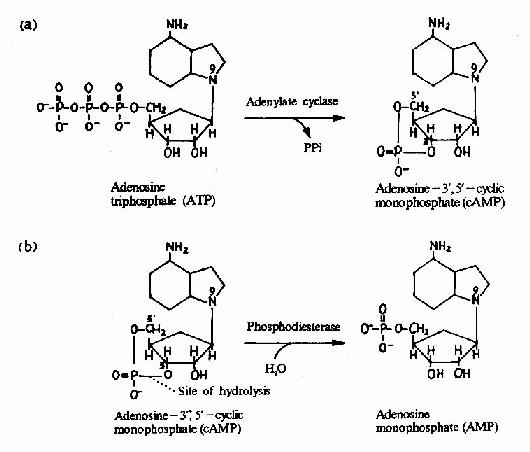
ͼ21��12��cAMP��������ˮ��
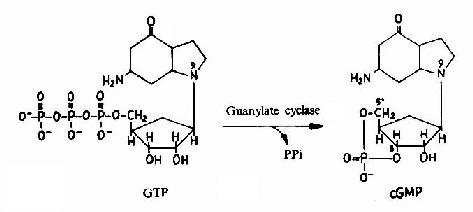
ͼ21��13��cGMP������

ͼ21��14��DAG��IP3������
����ϸ������ʹһ��������������ص㣺(1)��ΪС���ӣ��Ҳ�λ��������л;�������ģ�(2)��ϸ���е�Ũ�Ȼ�ֲ�����Ѹ�ٵظı䣻(3)��Ϊ�乹ЧӦ������������Ӧ�İз��ӣ���֪�İз�����ҪΪ���ֵ���ø��
��21��3��G����α�ǻ�����ЧӦ���Ӫ�
Ca���� ЧӦ���� ϸ������ʹ �з��� as �����ỷ��ø���Լ�↑ cAMP↑ ����øA����↑ ai �����ỷ��ø����↓ cAMP↓ ����øA����↓ aq ��֬øC����↑ Ca2+IP↑3DAG ����øC�↑ at cGMP��֬����ø↑���� cGMP↓ Na+ͨ���ر�
����cAMP�ǵ�һ�������ֵ�ϸ������ʹ���������ɵ������ỷ��øΪһ��Ҫ��Gαi��Gαs��ЧӦ���ӡ� cAMP�Ǻܶ༤�ص�ϸ������ʹ����һ����Ҫ��ϸ������ʹ������֬��������������֬øC�����£���PIP2(��������֬������)ˮ�����ɵ������ἡ�� (IP3)���Ͷ���(DAG)��
������Ҫָ�����ǣ���G����ż�������������ź�ת����������ϸ������ʹ��Ϊ�źŵĴ������⣬ϸ���ڻ��������������ź�ת����ʽ���ص�ϸ������ʹ����ʮ���������Խ��Խ�����С����������Ϊϸ������ʹ����ϸ�����ܵ��صĹ��̵��Բ�����
����G����ż����������G���鵼�µ��źŴ��ݹ��̿ɼ�ͼ21��15��ͼ21-16��
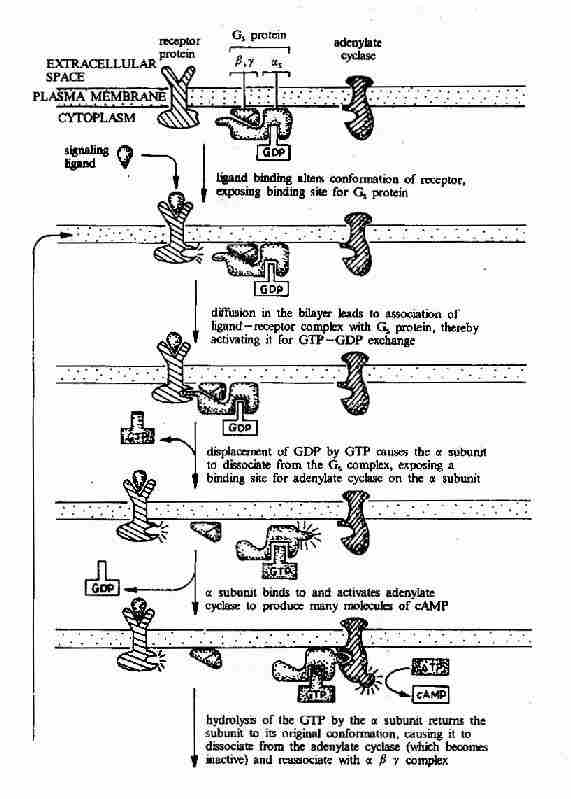
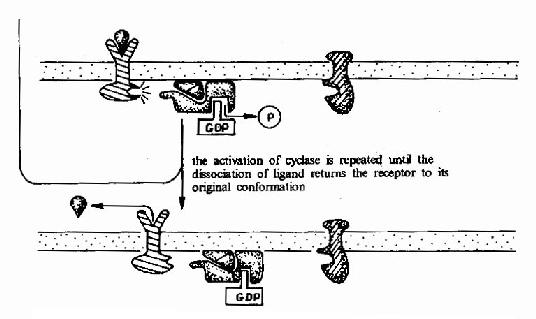
ͼ21��15��G����ż�������壭G���ף������ỷ��ø�ź�ת��;��ʾ��ͼ
ͼ21��16��G����ż�������壭G���ף���֬øC�ź�ת��;��ʾ��ͼ
����3.ϸ������ʹ���õ���Ҫ�з���
�������Gα����������Ӧ��ЧӦ���ӣ��Ӷ�ʹ��Ӧ��ϸ������ʹŨ�ȷ���Ѹ�ٵĸı䡣��Щϸ���ڵ���ʹ�ɷֱ���������Ӧ�İз��ӣ��Ӷ�ʹ��ϸ���еĸ���ø��͵����ӵĻ��Է����ı䡣��Щϸ������ʹ�����õİз�����ҪΪ���ֵ���ø(��21-3)
����(1)����øA
����cAMP������cAMP�����Ե���ø(cAMP��dependent Protein Kinase,���ΪcAPK),���Ϊ����øA(Protein Kinase A, PKA)��Ŀǰ��һ��������Ϊ���ϡ�cAMP������Ϊ�ü�ø�ı乹�������ʹ���Եĵ���øAת��Ϊ�л��Եĵ���øA(�����ʮһ�£�ͼ11��4)�� ��˵ĵ���øA�������ڶ�������֬��л��ص�ø�ࡢһЩ����ͨ����ijЩת�����ӣ�ʹ���Ƿ������ữ���ı������״̬��
����(2)����øG
����ϸ���ڵ���һ�ֻ���������ʹΪ�������ᣭcGMP��cGMP������cGMP�����Ե���ø ��cGMP��dependent Protein Kinase, cGPK)�����Ϊ����øG(Protein Kinase G��PKG)����PKAһ����PKG��Ŀǰ��Ϊ���ϵ�����������øG��cGMP��Ϊ�乹ЧӦ�������Ժ�ƽ�����к����Ϸḻ��1997�꣬���Ƿ� �֣�PKG�Ļ���ͻ�����Ӭ����ʳ��Ϊ�йء����ǿ����Ʋ⣬PKG���п�������ϵͳ���źŴ��ݹ����о�����Ҫ���á�
����(3)����øC
��������һЩ��Ҫ��ϸ������ʹ��������֬����������������DAG��PIP3(��������֬������)����֬����������������֬���������Լ�Ca2���ȵȡ���ЩС������ʹ��һ����Ҫ�з����ǵ���øC(Protein Kinase C��PKC)��
����PKC�ж���ͬ��ø��ʽ������ϣ����ĸ���У���PKCα��PKCβI��PKCβ���PKCγ�ȵȡ���ͬ��ͬ��ø�ڽṹ����֯�ֲ��ϸ��в�ͬ����Ը�������(��������ϸ����С������ʹ)���������в�𣬲��ҶԵ�����ѡ���ԡ�
����PKC��ϸ���������ֻ��ĵ����м���������ϸ�������Ͼ��йؼ��Եĵ������ã���һ��dz���Ҫ���ź�ת�����ӡ� ϸ���ź�ת�������еĶ���;�������Ե���PKC�Ļ���Ѿ��кܶ�ʵ���о�֤����PKC�����Ƽ�����ʹϸ��ʧȥ�������ֻ��̼��źŵķ�Ӧ��������Щ���ܶ� ������PKC�ĵ��ء����磬�ʴ�ϸ�����ѿ�����Ӧ(�ͷų�������֯����Ѫ�ܻ�������)�ǻ����̬��Ӧ����Ҫ��ʽ֮һ������ʵ������������PKC���Ƽ�Ԥ �ȴ���ϸ����ϸ���Ͳ���Դ̼��ź��ٷ����ѿ�����Ӧ������øı�ϸ��Ĥͨ�Եķ�ʽʹ���ڵ�PKC©����ϸ��Ҳ��ʧȥ�����ѿ�����Ӧ����������ʱ���ټ��� PKCʹ֮�ص�ϸ���У����ֿɻָ�ϸ�����ѿ�����Ӧ�������ܶ����Ƶ�ʵ���������ϸ���ĺܶ���������Ҳ�ܵ�PKC�ĵ��ء�
����(��)���ο�Ĥ���弰���ź�ת��
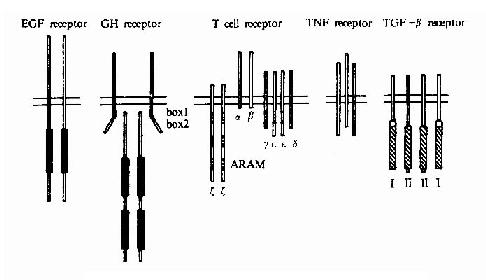
���������������Ӻ�ϸ�����ӵ�����Ϊһ��ṹ��Ϊ���ο�Ĥ���ǵ��ס����ߴο�Ĥ����(G����ż��������)���Ӧ�������Ϊ���ο�Ĥ���壬�����ǵĿ�Ĥ����Ϊ����һ���Եģ��������ߴο�Ĥ���������з����Ŀ�Ĥ���Ρ�
�������ο�Ĥ����������ṹ�ص�ɽ�һ���ֳɶ��������Ǽ��壬��������21��4��ͼ21-7����
ͼ21��17������ο�Ĥ����Ĵ����Ծ���
Table21-4 Families of Receptors Activated by Dimerization or Oligomerization
Receptor Type Family Examples Characteristics ����
�� PDGF receptor PDGFR-α,PDGFR-βSCFR Five immunoglobulin-like family (Kit),CSF-R(Fms),Flk-2 domains extracellularly EGF receptor family EGFR(ErbB),ErbB2(Neu),ErbB3,ErbB4 Two cysteine-rich IGF receptor family insulin R,IGF-1R domains extracllularly HGF receptor family HGFR(Met),MSPR(Ron) Disulphide-bound heteroteramer of a and βchain VEGF receptor family Flt-1,Flk-1(KDR) Seven immunolobulin-like domains extracellularly Neurotrophin receptor family Trk,TrkB,TrkC �� Eph receptor family Eph ,E1k,Eck,Cck5,Sek,Erk Two FNIII-like domains and a cysteinerichdomain extracellularly ����
�� Class 1 cytokine receptor family �� CH
receptor subfamily GHR,EPOR,PRLR,G-CSFR Form homodimers IL-3 receptor subfamily IL-3R,GM-CSFR,IL-5R Form complexes with the cβ subunit IL-6 receptor subfamily IL-6R,LIFR,CNTFR,IL-11R Form complexs with gp 130 IL-2receptor subfamily IL-2R,IL-2Rβ,IL-4R,IL-7R Form complexes with il-2Rγ �� IFN-α/βR,IFN-γRα,IFN-γRβIL-10R �� TNF receptor family ���� TNFR-1,TNFR-11MLNGFR,CD40,OX-40,Fas,CD27,CD30 Form trimers �� TCR Complex of α,β,γ,δ,ε,ζandηsubunits �� BCR Complex of IgM and heterodimers ofα/βsubunits Serine/threonine kinase receptor family Type �� receptor family TGFβR-��,ActR-��,ActR-��B Form hetero-oligomers with type Ireceptors,i, e,TGFβ-I,ActR-1,ActR-1B,BMPR-1A,BMPR-1B,ALK-1
����Receptor families and subfamilies discussed in the text are presented .Abbreviations used: R, receptor; R, receptor; PDGF,platelet-derived growth factor; SCF, stem cell factor; CSF, colony-stimulationg factor; EGF,epidermal growth factor; FGF, fibroblast growth factor; IGF, insulin-like growth factor;HGF, hepatocyte growth factor;MSP,macrophage-stimulating protein;VEGF,vascular endothelial growth factor;FN,fibronectin;GH,growth hormone;EPO,erythropoietin;PRL,prolactin;IL,interleukin;LIF,leukemia inhibitory factor;CNTR,ciliary neurotrophic factore;IFN,interferon;TNF,tumor necrosis factor;Lngfr,low affinity nerve growth factor rcecptor;TCR,T ,cell rcecptor;BCR,B cell rcecptor;receptor;TGFR,transfoming growth factor βAct,activin;BMP,bone morphogenic protein,Alternative desingations are given within parentheses.
�������ο�Ĥ�������鵼���źŴ�����ת��������G����ż��������鵼���ź�ת�����źܴ��������Ѿ�֪����G�� ��ż�����������鵼����Ҫ�Ǿ���G���ļ��Ȼ����������Ӧ��ЧӦ���ӣ�����������Ҫ���ǵ���ϸ������ʹ�������ֲ���Ѹ�ٸı�Ӷ����ڰз��ӵĻ��Բ��� ��ϸ���Ĺ���״̬�����ο�Ĥ����鵼���ź�ת����������Ҫ�ǵ����ӵ�����ã������е����Ұ��ἤø�Ĺ㷺���롣����Щ�ź�ת��;�����˽��ھ�ʮ����� ȡ����������Ҫ�Ľ�չ��Ϊ���ٺ�������Щ�ź�ת�����̣�����������Ҫ֪��������һ���̵���Ҫ�ź�ת�����Ӻ����е�һЩ����ṹ��
����1.������Ҫ�Ľ鵼���ο�Ĥ�����ź�ת���ĵ����ӵĽṹ������
����(1)����ø
��������ø��ָ�ܹ���γ�������������ṩ�������ת�������ﵰ�İ����������ϵ�һ����ø�����ṩ������� ATP��Ҳ����������������������ᡣ���ڵ���ø�����Ƕ����ģ���˵���ø�Ǹ��ݵ����а�����л��������Զ����Ǹ��ݵ��ﵰ��������������ġ��� ������ѧ������ίԱ�Ὠ�齫����ø��Ϊ����ࣺ
������ ø ������ŵ����� ����˿���ᣯ�հ��ἤø ˿���ᣯ�հ����ǻ� �����Ұ��ἤø �Ұ���ķ��ǻ� �����飯���������ἤø �����һ���ε������ ���װ��װ��ἤø �ϻ� �����춬���ᣯ�Ȱ��ἤø ����
����ǰ���༤øĿǰ�˽�Ľ϶࣬�ܶ��Ѿ������cDNA��¡�����Ժ����ߵ��˽��Ժ����ޡ�
��������˿/�հ��ἤø�͵����Ұ��ἤø�ķ������21��5����
��21-5������ø���ત
1.����˿����/�հ��ἤø ����(1) Cyclic nucleotide regulated ����A. PKA α cAMP��dependent protein kinase catalytic,α form ������B. PKA βcAMP��dependent protein kinase catalytic, β form ������C. PKA γcAMP��dependent protein kinase catalytic,γ form ������D. cGPK��α cGMP��dependent protein kinase α form ������E. cGPK��β cGMP��dependent protein kinase β form ����(2) Diacylglycerol regulated:[BH] ������PKC α protein kinase C α ������PKC β protein kinase C β ������PKC γ protein kinase C γ ������PKC δ protein kinase C δ ������PKC ε protein kinase C ε ������PKC ζ protein kinase C ζ ������PKC η protein kinase C η ����(3)Calcium/calmodulin regulated[BH] ����CaM�� ��Calcium/calmodulin��dependent protein kinase �� ������CaM��α ��Calcium/calmodulin��dependent protein kinase ��α ������CaM��β ��Calcium/calmodulin��dependent protein kinase ��β���� ������CaM��γ ��Calcium/calmodulin��dependent protein kinase ��γ ����CaM��Calcium/calmodulin��dependent protein kinase �� ������CaM��Calcium/calmodulin��dependent protein kinase �� ���������� ��PSKHI Putative protein��serine kinase ����(4) Ribosomal S6 protein kinase ���������� ��S6k��,S6K��,70kDaS6 ����(5) Serpentine receptor kinase ���������� ���� β ARK β��Adrenergic receptor protein kinase ���������� ���� β ARK��related protein kinase ���������� ���� Rhodopsin kinase ����(6) Casein kinase �� ����(7) Glycogen synthase kinase 3 ����(8) cdc 2 family ����(9) cdc 2 related protein kinase ����(10)MAP kinase (mitogen activated protein kinases) ���������� ��EPK��1, ERK��2, ERK��3, ERK��5(extracellular signal��regulated kinase) ���������� ��JNK(c��Jun N��terminal kinase), P38, etc. ����(11)MOS/Raf protein kinase 2.�����Ұ��ἤø ����(1) Growth factor receptors ���������� ��EGFR: EGF
R.erbB2,erbB3 ���������� ��INSR:INSR, IGF��1R ���������� ��FGFR:FGFR, FGFR2, c��kit ���������� ��DGFR:PDGFR, CSFIR ����(2) Non��receptor tyrosine kinase ���������� ��src family: src,Yes,Lyn,Fyn,Lck,etc ���������� ��Syk family:Syk, ZAP70 ���������� ��JAK family: JAK1,JAK2, JAK3 ���������� ��Tec family: Btk,Itk,etc
���������г��Ķ��ֵ���ø��������Ҫ���ź�ת�����ӣ��ڴ˲�����һһ���ܡ�����Щ����ø�У�����ϸ������Ӱ��ϴ����PKA��PKG��PKC��MAPK��PTK�ȡ�PKA��PKG��PKC����G����ż���������ἰ��������������MAPK��PTK��
������MAP��ø(Mitogen Activated Protein Kinase, MAPK)
����MAPK���ڵ���˿/�հ��ἤø���ǽ���Ĥ����ת���봫�ݵ��źŲ��������ϸ�����ڵ�һ����Ҫ���ӣ��ڶ����� ���źŴ���;���о����йؼ������á���δ�ܴ̼���ϸ���ڣ�MAPKΪ��ֹ�ͣ�����������η��ӗ�MAPKK(MAp Kinase Kinase)�����ữ�����źź�MAPK�����ڵ��հ�����Ұ���������ữ���Ӷ���Ϊ���ʽ��MAPK(ͼ21-18)��Ŀǰ��MAPK�����δ��� ��һ��
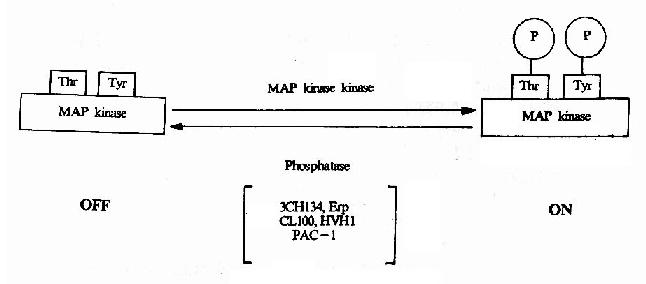
ͼ21-18��MAPK�Ļ
ͼ21-19��MAPK������
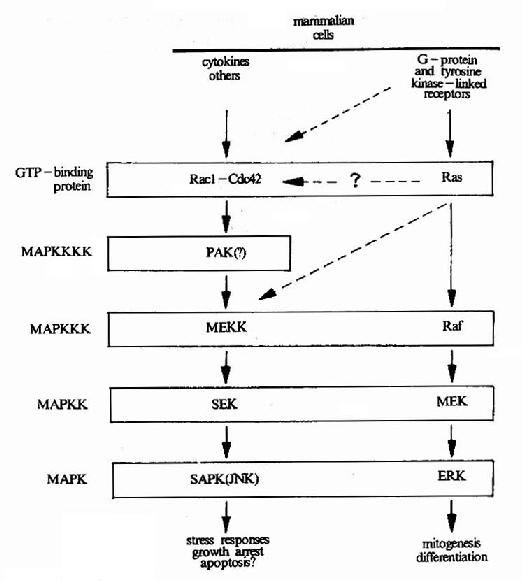
�����ɵ���ø���ɵ�MAPK����ϵͳ������ʹMAPK���ữ��MAPKK���ܵ�MAPKKK(MAp Kinase��Kinase Kinase)�ĵ��ڡ���������ϵͳ�����������η��ӵĵ���(ͼ21-19)��
����MAPK�������Ժ�ת����ϸ�����ڡ��ں��ڣ�������ʹһЩת¼���ӷ������ữ�Ӷ��ı���ڻ�������״̬�� ���⣬��Ҳ����ʹһЩ������ø�������ữʹ֮���Է����ı䡣Ŀǰ�Ѿ�֪����MAPK�������ϸ�����ܵĵ��أ���������ϸ����ֳ���ֻ������������У������� �ؼ������á�
����MAPK;���ڹ�Ӭ���ѷ�����7�ֲ������ͣ��ڲ��鶯��ϸ������֤ʵ����������MAPK��Ա����ERK��JNK��P38�����ǵĵ��ػ��ƺ����õİз��Ӿ��������졣
�����ڵ����Ұ��ἤø(Protein Tyrosine Kinase, PTK)
���������Ұ��ἤø�����ڵ������е��Ұ���л�ʹ֮���ữ���ܶ�ϸ���ź�ת�����������¼�Ϊ���ֵ����ʵ��Ұ����� �ữ����ϸ����������ֻ������У��Ұ������ữ�־�������������ã������������������ú�����ϸ������ֳ����������ϸ������ֳ������Tϸ����Bϸ���� �ʴ�ϸ���Ļ��������˲�䷢���Ķ��ֵ����ӵ��Ұ������ữ�������Ұ��ἤø�����Ƽ������������ϸ����Ӧ��Ӧ��
�������ݵ����Ұ��ἤø��ϸ���ڵ�λ�ÿ��Խ����Ϊ���ࣺ
����A.�����Ұ��ἤø����
������һ�൰���Ұ��ἤøΪ��Ĥ���ף�����ⲿ��Ϊ�����������м��п�Ĥ�������ڲ��ֺ��е����Ұ��ἤø�Ĵ��ṹ��(ͼ21��17)��
���������Ұ��ἤø�����������Ϻ������γɶ����壬�̶�����ø���Ե����ߣ�ʹ������ڲ��ֵ��Ұ������ữ��ǿ�����ữ������ø���Խ�һ����ǿ���������Ҫ���ǣ����ữ���������Ľ������SH2�ṹ��(����)���źŷ��ӣ��Ӷ����źŴ��������η��ӡ�
����B.λ�ڰ������ֵĵ����Ұ��ἤø
�����������ຬ�ж���PTKs����Щ����ø��������崫�ݵļ����źţ����������������ĵ��ڶ��������Ըı䣬����Ҫ��������Ϊ���������ЧӦ����֮����м�鵼���ӡ���֪����Ҫ�����У�
����(1)src���壺src��Fyn��Lck��Lyn�ȣ��������ϴ��ڣ��������������Ϻ��
����(2)Tec���壺Btk��Itk��Tec�ȣ��������ϻ�ϴ��ڣ������Ϻ��
����(3)ZAP70���壺ZAP70��Syk�������ữ�������Ϻ��
����(4)JAK���壺JAK1��JAK2��JAK3�ȡ�
������ЩPTKs����ֱ���������γɸ��������ӵ����α�����ڴ��������źŹ��������Ž����������á����ǵĽṹ��ͼ21-20��
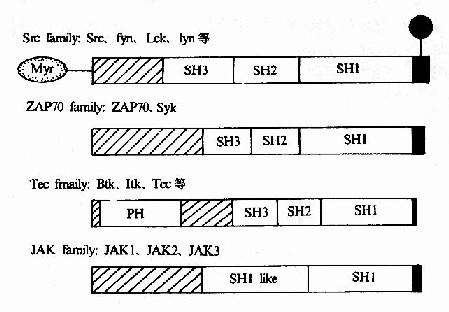
ͼ21��20������PTK�Ľṹ
����C.���ڵ����Ұ��ἤø
�����ֵ��Ұ��ᵰ��øλ�ڰ�Ĥ�ϻ�����ڣ�������ȴ���ֺ���Ҳ�������Ұ��ᵰ��ø��������ź��ں��ڵĴ� ������Ҫ���塣��Ҫ�ĺ���PTKs��Abl��Wee��Abl�ȴ����ڰ����ڣ�Ҳ�����ڰ����У��ѷ���������ת¼���̺�ϸ�����ڵĵ��ڣ�Weeֻ�����ں� �ڣ����ɵ���Cyclin-2�Ļ��ԣ����������ữ����ϸ��������˿�����ھ��е������á�
����(2)��������ø(Protein Phosphotase)
������������(��)ø��ָ���д��Ѿ����ữ�ĵ����ӷ���ȥ���ữ��Ӧ��һ��ø���ӡ������뵰��ø���Ӧ�� �ڣ���ͬ���������ữ��ȥ���ữ��һ��Ҫ�ĵ����ʻ��ԵĿ���ϵͳ�����絰��ø�������ữ���ڵ��ﲻͬ���еı���Ϊ�������ߣ��еı���Ϊ���Խ���һ���� ��������ø������ȥ���ữ��ӦҲ���ڲ�ͬ�ĵ����в�ͬ�ķ�Ӧ(ͼ21-21)��
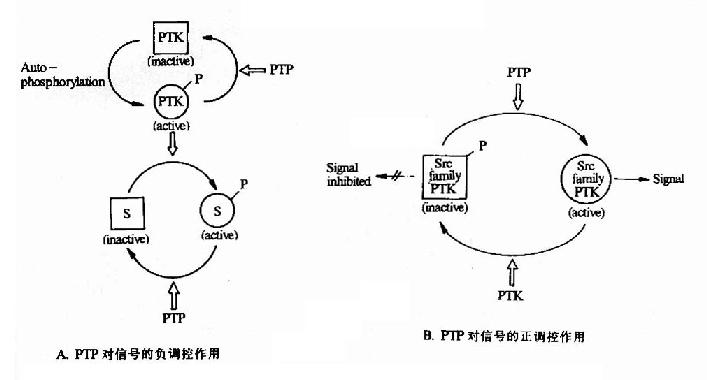
ͼ21-21�������Ұ�������ø�Ե����Ұ��ἤø�ĵ������ê�
PTK��Protein Tyrosine Kinase�� PTP��Protein Tyrosine Phosphotase S:Substrate
������������ø�ķ���Ҳ�뵰��ø���ƣ��Ǹ����������õİ�����л������ġ�Ŀǰ��֪�ĵ�������ø��������˿����/�հ�������ø�͵����Ұ�������ø�����࣬����и���ĵ�������ø����˫�����ã�����ͬʱ�������Ұ����˿����л���
��������ø�͵�������ø�����������ĵ�����ǵĴ����õ������Լ�����ϸ���ڵķֲ������Ծ������ź�ת��;���ľ�ȷ�ԡ�
����(3)�ͷ�����G����(Small G Protein)
����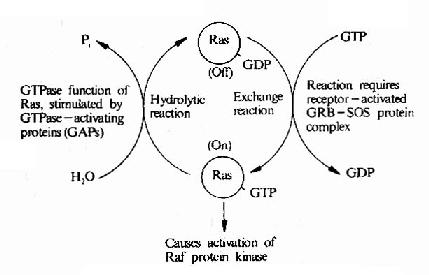
ͼ21-22��Ras�Ļ�����������
�����ͷ�����GTP��ϵ����ڶ���ϸ����Ӧ�о��п������ã�����λ��MAPKϵͳ�����Σ���һ����Ҫ���ź�ת�����ӡ���һ�������ֵĵͷ�����G������Ras�������һ�൰���ʱ���ΪRas�������Ա���������Ǿ���һ��GTPø�ṹ�ɣ��ཫ֮��ΪRas��GTPø��
�����ͷ�����G���Ĺ�ͬ�ص��ǣ��������GTPʱ����Ϊ���ʽ����ʱ�����������η���ʹ֮�������GTPˮ���ΪGDPʱ(����ΪGTPø)��ظ����ǻ״̬����һ����Gα���ƣ�����Ras����ķ��������Ե���Gα��
������ϸ���д�����һЩר�ſ��Ƶͷ�����G�����Եĵͷ�����G���������ӣ���Щ����������Ĥ�����źŵ�Ӱ�죬 ���ڵͷ�����G�����ԣ��ͷ�����G������������MAPKϵͳ������Щ���������У��еĿ�����ǿ�ͷ�����G���Ļ��ԣ��������ʺ����ύ������(GEF) �������ʺ����������������(GDI)���еĿ��Խ��͵ͷ�����G�����ԣ���GTPø�����(GAP)��(ͼ21��22)��
����Ŀǰ��֪��Ras�������Ա�ѳ���50�֣������ֿ��Է�Ϊ�����Ǽ��壬��ϸ���ڷֱ���Ʋ�ͬ���ź�ת��;����ÿһ����Ա��������Ե��ص㡣
����(4)�鵼�ź�ת����������õĽṹԪ��
����ϸ���д������ڶ���ź�ת�����ӣ�����������ʶ������ö����ɲ�ͬ��ϸ��ת��;������?90����� �����������������ź�ת�������д�����һЩ����Ľṹ����Щ�ṹ���Լ��50��100�������ṹ�ɣ������ڲ�ͬ���ź�ת�������о��кܸߵ�ͬԴ�ԡ� ��Щ�ṹ�����������ϸ���н鵼�ź�ת�����ӵ��ʶ�𣬹�ͬ�γɲ�ͬ���źŴ��������Ϊ�ź�ת��;��(Signal Transduction Pathway)���������γ��ź�ת��������(Signal Transduction Network)�����仰������Щ�ṹ�����·�еĽ�ͷԪ��һ���Ѳ�ͬ���źŷ���������������Щ�ṹ��Ϊ���ؽ��Ԫ��(Modular Binding Domain)��Ŀǰ�Ѿ�֪���˽�ʮ�������Ľṹ�����磺
����SH2 domain (src Homology 2 domain)SH3 domain��src Homology 3 domain)
����PH domain��(Pleckstrin Homology domain)
����PTB domain(Protein Tyrosine Binding domain)
����A.SH2�ṹ����Լ100����������ɣ��鵼�źŷ����뺬�����Ұ��ᵰ���ӵĽ�ϡ����ֽ���������Ұ���л������ữ������Χ�İ�����л������ɵĻ���(motif)��
��������YEEI��src family��SH2���
����YMXM��growth factor receptor��P13Kp85��SH2���
����YVIP��PLC�SH2���
����YXNX��Grb2��SH2���
����B.SH3�ṹ����50��100����������ɣ��鵼�źŷ����븻��������ĵ����ӵĽ�ϣ����������븬����л����ڽ�������л������ɵĻ���������ء�
��������RKLPPRPSK��P13K������Ϊ9.1μM
����PALPPLPRY��P13K������Ϊ17μM
����C.PH�ṹ����100��120����������ɣ��书����δ��ȫȷ����Ŀǰ��֪����������֬�����PIP2��PIP3��IP3�Ƚ�ϡ�ͬʱҲ���֣�һЩ�����ӣ���PKC��G����βγ�ǵ�λҲ������PH�ṹ���ϡ�
����D.PTB�ṹ����Լ160����������ɣ���SH2һ����PTB�ṹ��Ҳ����ʶ��һЩ�������Ұ���Ļ������ϻ�����SH2�ṹ���������
������Ϊ���ؽ��Ԫ���������ڽṹ�������������ص㣺
������һ���źŷ��ӿ��Ժ����������ϵĵ��ؽ��Ԫ��(ͼ21-23)����˿���ͬʱ���������ϵ������źŷ������ϣ����磬�ڵ����Ұ��ἤøBtk�м���PH�ṹ��SH3�ṹ���SH2�ṹ���3�����ؽ��Ԫ����

ͼ21-23���ź�ת�������еĵ��ؽ��Ԫ����
����Y-Kinase��tyrosine kinaseDBD:DNA binding domain��PP:Pro��rich
����ABD:actin��binding domainGAP:��GTPase��activating domain
������ͬһ����ؽ��Ԫ���ɴ����ڶ��ֲ�ͬ���ź�ת�������У����磬PH�ṹ�������ijЩ����ø���ͷ�����G�� ���ڷ��Ӽ�ϸ���Ǽܵ��ȶ����ź�ת�������С���Щ���ؽ��Ԫ����һ���ṹ��Ȼ�Dz�ͬ�ģ���˶�����ϵ��źŷ��Ӿ���ѡ���ԣ����DZ�֤�źŷ�������� ���������ԵĻ�����
��������Щ�ṹ������Ϊ�Ǵ��ṹ��
����2.�������͵��ź�ת��;��
����(1)��Ƥ�������������鵼���ź�ת��;��
������Ƥ���������������壭��Ƥ�������������Ϻ������һϵ��ϸ���ڱ仯������ʹϸ�������ֻ�����ֳ����Ƥ���� ����������һ�������Ұ��ᵰ��ø���������Ұ��ᵰ��ø→Ras→MAPK����;���DZ�Ƥ�������Ӵ̼��źŴ��ݵ�ϸ�����ڵ�����Ҫ;�����������³�Ա�� �ɣ���Ƥ������������→����SH2�ṹ��Ľ�ͷ����(��Grb2)→�����ʺ������ͷ�����(��SOS)→Ras����→MAPKKK(�� Raf1)→MAPKK→MAPK→ת¼���ӵ�(ͼ21��24)��
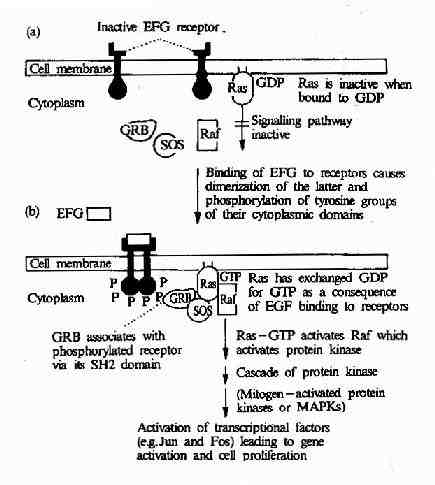
ͼ21-24��EGF����鵼���ź�ת������
������Ƥ���������������Ϻ���ʹ���巢�������廯���Ӷ��ı�������Ĺ���ʹ���еĵ����Ұ��ἤø������ ǿ�������������Ұ���л��������ữ�����ữ��������γ����뺬SH2�ṹ��ĵ�����Grb2��ϵ�λ�㣬����Grb2������Ľ�ϡ�Grb2�������� SH3�ṹ�ò�λ��һ�ֳ�ΪSOS�������ύ�����ӽ�ϣ�ʹ֮���Ըı䣬SOS���һ���Ras�������Ras������MAPK����ϵͳ������ERK �ļ�����ERKת�Ƶ�ϸ�����ڣ�����ijЩת¼���ӵĻ��Ըı�Ӷ��ı����ı���״̬��ϸ������ֳ��ֻ����̡�
����(2)γ������������鵼���ź�ת��
����γ�����������ɻTϸ�������ģ������дٽ���ԭ��ʺ�����������ʶ������ã����ɴٽ�Bϸ������������γ�� ���������������Ժ�Ҳ���Ե�����������廯�������廯��������Լ���JAK��STATϵͳ�����߽������ش̼��źŴ�����ڡ�JAK(Janus Kinase)Ϊһ�ִ����ڰ����еĵ����Ұ��ἤø��������ʹ�������������ữ��STAT(Signal Transducerand Activator of Transcription)����ͨ����SH2�ṹ��ʶ�����ữ�����岢��֮��ϡ�Ȼ��STAT��������Ұ�������ữ���Ұ������ữ��STAT����� ���γ��л��Ե�ת¼���ӣ�Ӱ�����ı���(ͼ21��25)��

ͼ21-25��γ������������鵼���ź�ת������
����JAK��Janus kinase
����STAT: Signal Transducer and Activator of Transcription
����GAS: �interferon��activated sequence element
�������������ź�ת��;�������Ƕ����ź�ת��;���Ĵ���������90���������ѧ������ϸ���ź�ת���ķ��ӻ����о������Ѿ�ȡ����һЩ�ɾͣ����������ϸ���д��ڵ�ȫ����������ϵͳ��ʮ��ңԶ���д���ѧ���Dz���Ŭ�������¸�����ʵ��������ʶ���ҵ�Ը����
��һƪ�����������ְ�����
��һƪ��ϸ����������ź�ת������










